

Patterned activity via spinal dorsal quadrant inputs is necessary for the formation of organized somatosensory maps
- PMID: 14614091
- PMCID: PMC6741022
- DOI: 10.1523/JNEUROSCI.23-32-10321.2003
Patterned activity via spinal dorsal quadrant inputs is necessary for the formation of organized somatosensory maps
Abstract
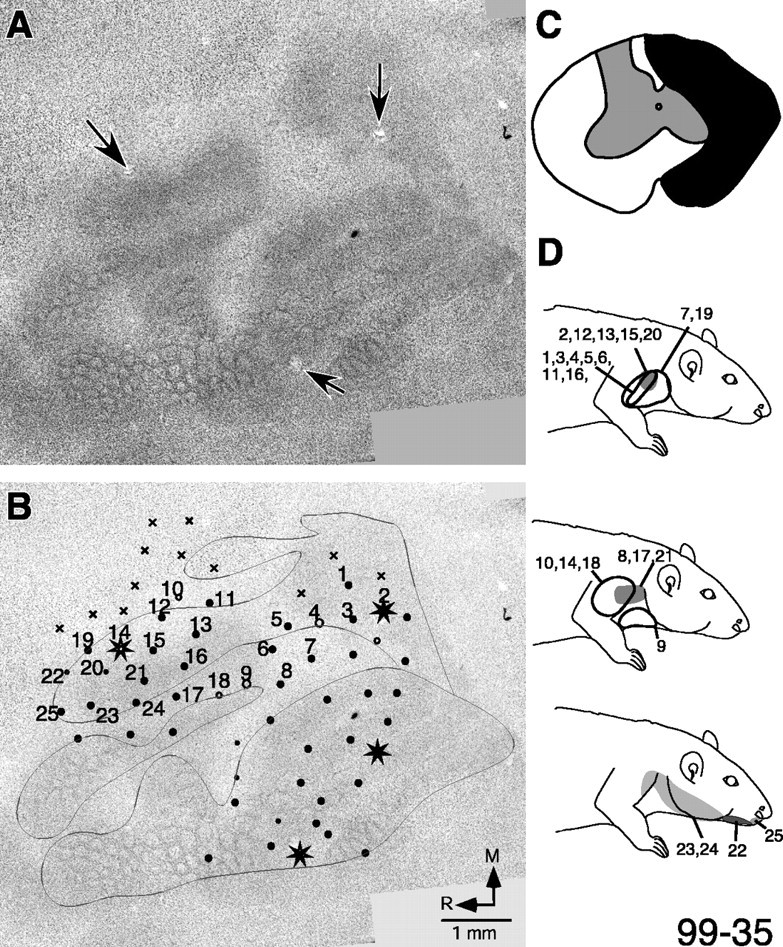
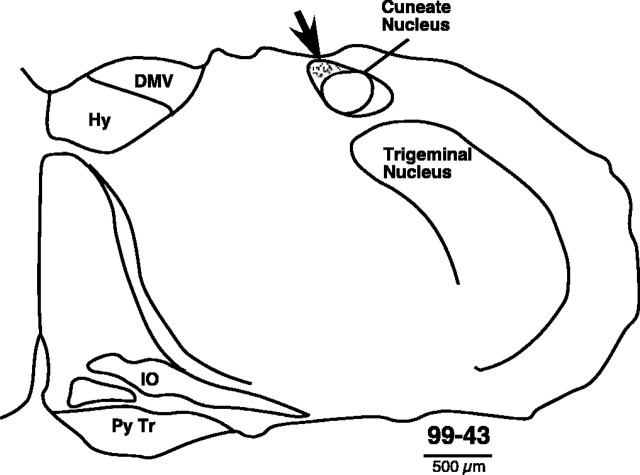
The normal development of the somatosensory system requires intact sensory inputs from the periphery during a critical window of time early in development. Here we determined how the removal of only part of the ascending spinal inputs early in development affects the anatomical and neurophysiological development of the somatosensory system. We performed spinal overhemisections in rat pups at C3/C4 levels on the third day after birth. This procedure hemisects the spinal cord on one side and transects the dorsal funiculus on the other side. When the rats were 6-8 months old, the responsiveness and somatotopy of the primary somatosensory cortex (S1) contralateral to the hemisection were determined using standard multiunit mapping techniques. Sections of the flattened cortex were processed for cytochrome oxidase activity, Nissl substance, or myelin. We found that histologically apparent modules that are normally present in the regions of the forepaw and the hindpaw representations were absent, whereas the lateral barrel field representing the face was completely normal. The neurons in the forepaw regions of S1 either did not respond to the stimulation of the skin of any region of the body or responded to the stimulation of the upper arm afferents that enter the spinal cord rostral to the site of the lesion. The results show that a lack of normal sensory inputs via ascending pathways in the dorsal spinal cord during early development results in massive anatomical and neurophysiological abnormalities in the cortex. Intact crossed spinothalamic pathways are unable to support the normal development of the forepaw barrels.
Figures






References
-
- Chapin JK, Lin C-S ( 1984) Mapping the body representation in the SI cortex of anaesthetized and awake animals. J Comp Neurol 229: 199-213. - PubMed
-
- Crockett DP, Maslany S, Harris SL, Egger MD ( 1993) Enhanced cytochrome-oxidase staining of the cuneate nucleus in the rat reveals a modifiable somatotopic map. Brain Res 612: 41-55. - PubMed
-
- Dado RJ, Katter JT, Giesler Jr GJ ( 1994) Spinothalamic and spinohypothalamic tract neurons in the cervical enlargement of rats. II. Responses to innocuous and noxious mechanical and thermal stimuli. J Neurophysiol 71: 981-1002. - PubMed
-
- Dawson DR, Killackey HP ( 1987) The organization and mutability of the forepaw and hind paw representations in the somatosensory cortex of neonatal rat. J Comp Neurol 256: 246-256. - PubMed
-
- Diener P ( 2002) Enriched environments improve skilled movements following neonatal cervical spinal cord injury in rats. Soc Neurosci Abstr 28: 633.620.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous