

Blockade of inflammation and airway hyperresponsiveness in immune-sensitized mice by dominant-negative phosphoinositide 3-kinase-TAT
- PMID: 14623911
- PMCID: PMC2194122
- DOI: 10.1084/jem.20030298
Blockade of inflammation and airway hyperresponsiveness in immune-sensitized mice by dominant-negative phosphoinositide 3-kinase-TAT
Abstract
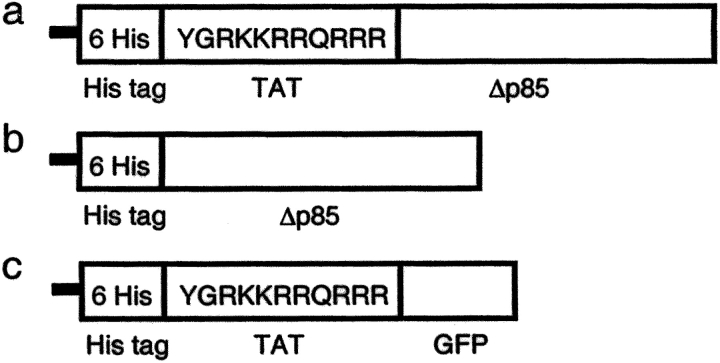
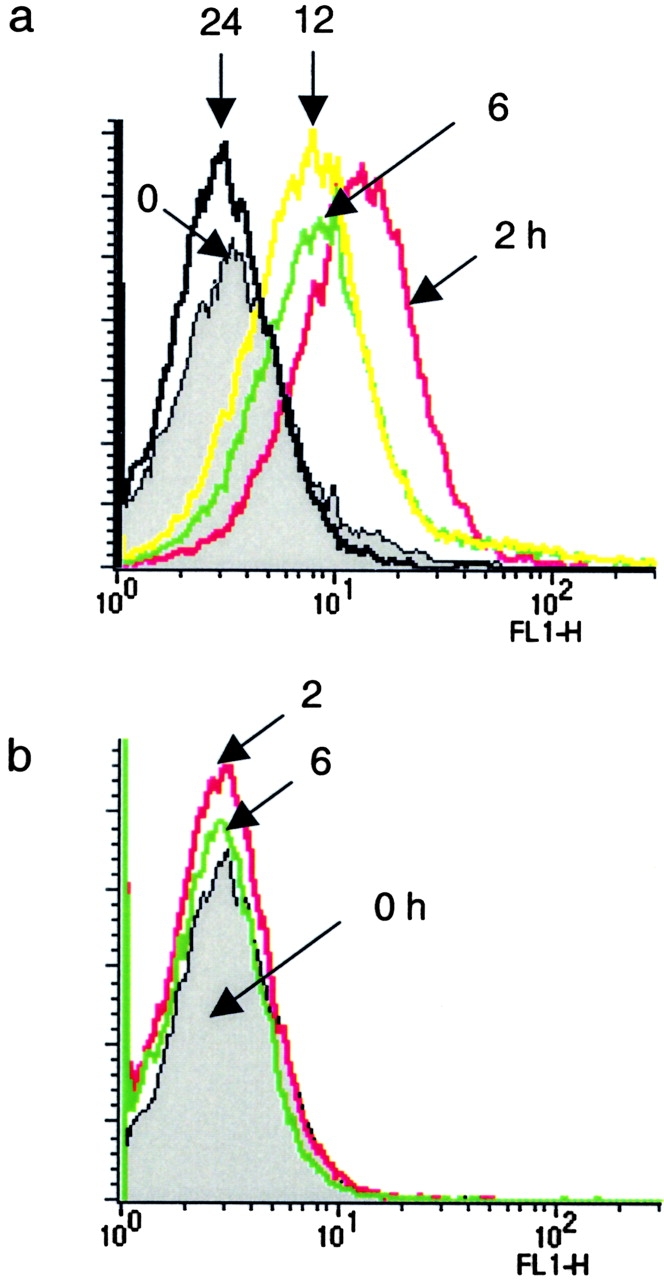
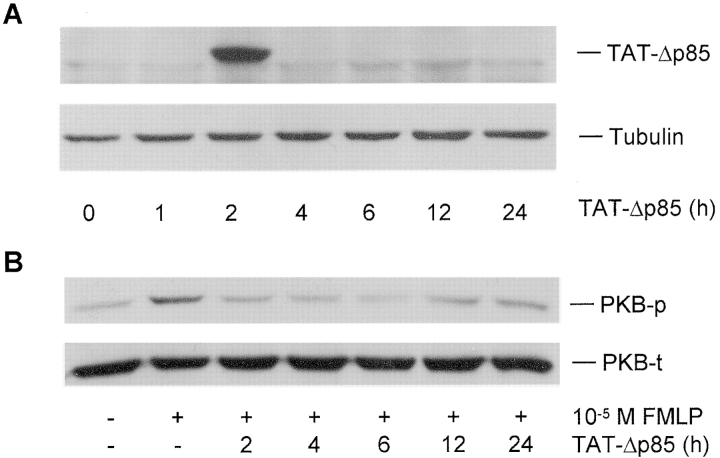
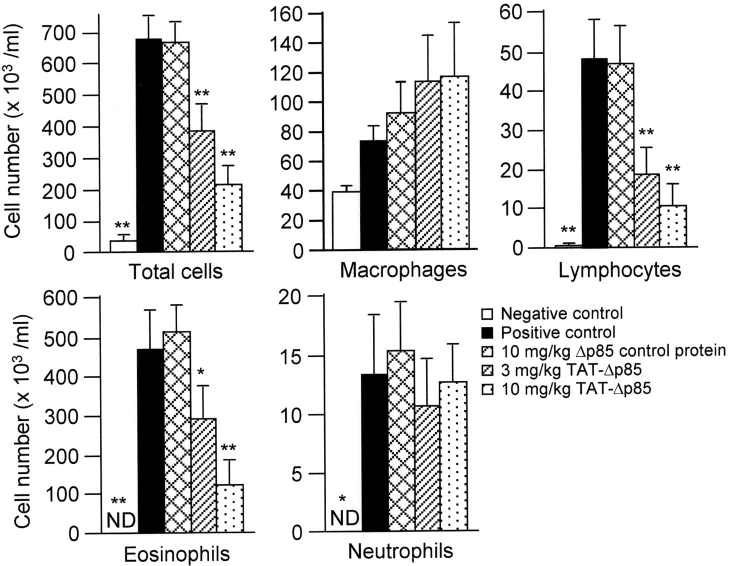
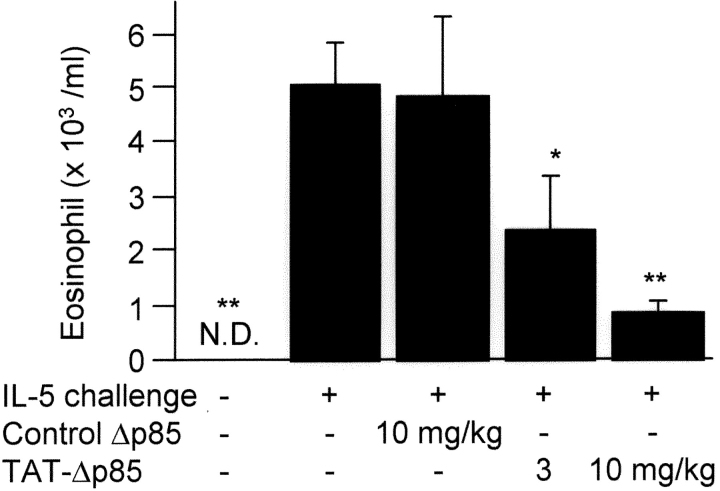
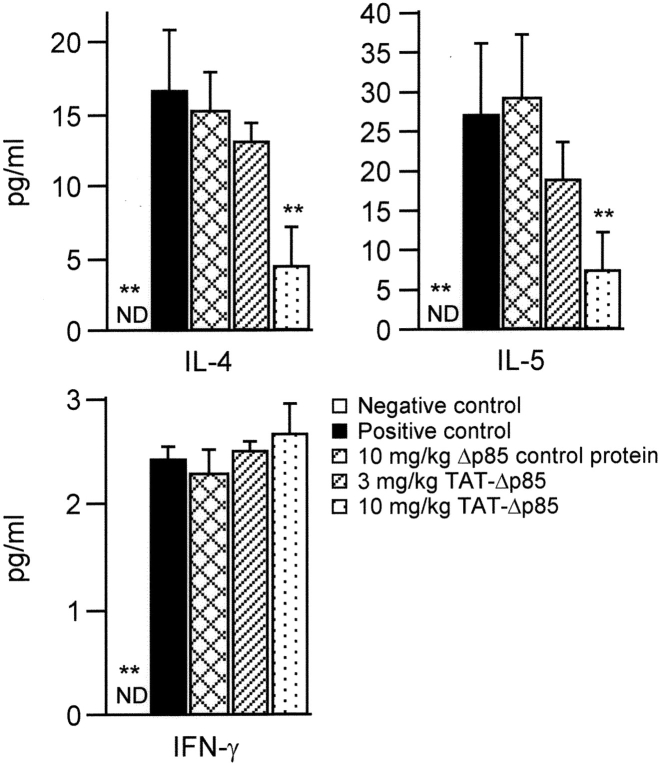
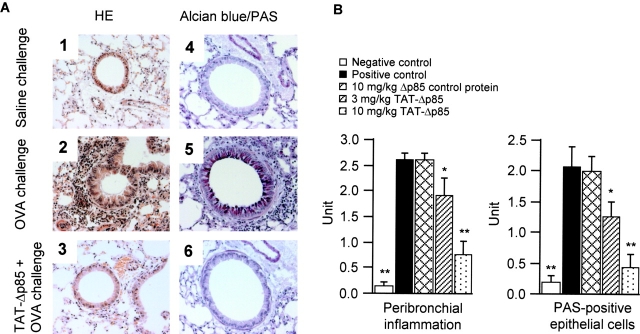
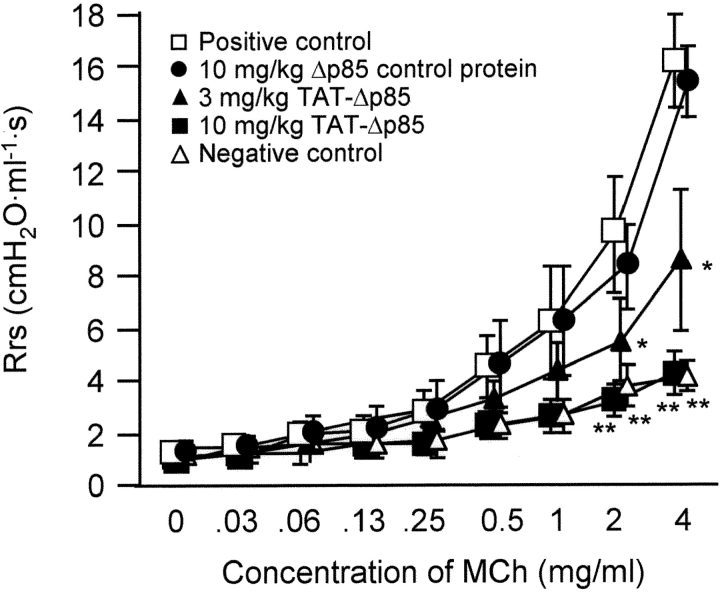
Phosphoinositide 3-kinase (PI3K) is thought to contribute to the pathogenesis of asthma by effecting the recruitment, activation, and apoptosis of inflammatory cells. We examined the role of class IA PI3K in antigen-induced airway inflammation and hyperresponsiveness by i.p. administration into mice of Deltap85 protein, a dominant negative form of the class IA PI3K regulatory subunit, p85alpha, which was fused to HIV-TAT (TAT-Deltap85). Intraperitoneal administration of TAT-Deltap85 caused time-dependent transduction into blood leukocytes, and inhibited activated phosphorylation of protein kinase B (PKB), a downstream target of PI3K, in lung tissues in mice receiving intranasal FMLP. Antigen challenge elicited pulmonary infiltration of lymphocytes, eosinophils and neutrophils, increase in mucus-containing epithelial cells, and airway hyperresponsiveness to methacholine. Except for modest airway neutrophilia, these effects all were blocked by treatment with 3-10 mg/kg of TAT-Deltap85. There was also significant reduction in IL-5 and IL-4 secretion into the BAL. Intranasal administration of IL-5 caused eosinophil migration into the airway lumen, which was attenuated by systemic pretreatment with TAT-Deltap85. We conclude that PI3K has a regulatory role in Th2-cell cytokine secretion, airway inflammation, and airway hyperresponsiveness in mice.
Figures
Similar articles
-
Regulation of interleukin-5-induced beta2-integrin adhesion of human eosinophils by phosphoinositide 3-kinase.Am J Respir Cell Mol Biol. 2005 Jul;33(1):65-70. doi: 10.1165/rcmb.2005-0076OC. Epub 2005 Mar 31. Am J Respir Cell Mol Biol. 2005. PMID: 15802551 Free PMC article.
-
Blockade of airway inflammation and hyperresponsiveness by HIV-TAT-dominant negative Ras.J Immunol. 2003 Oct 15;171(8):4379-84. doi: 10.4049/jimmunol.171.8.4379. J Immunol. 2003. PMID: 14530363
-
Activation of group IV cytosolic phospholipase A2 in human eosinophils by phosphoinositide 3-kinase through a mitogen-activated protein kinase-independent pathway.J Immunol. 2003 Oct 15;171(8):4399-405. doi: 10.4049/jimmunol.171.8.4399. J Immunol. 2003. PMID: 14530366
-
Phosphoinositide 3-kinase δ (PI3Kδ) in respiratory disease.Biochem Soc Trans. 2018 Apr 17;46(2):361-369. doi: 10.1042/BST20170467. Epub 2018 Mar 9. Biochem Soc Trans. 2018. PMID: 29523773 Review.
-
Phosphoinositide 3-Kinase in Asthma: Novel Roles and Therapeutic Approaches.Am J Respir Cell Mol Biol. 2017 Jun;56(6):700-707. doi: 10.1165/rcmb.2016-0308TR. Am J Respir Cell Mol Biol. 2017. PMID: 27977296 Free PMC article. Review.
Cited by
-
Unleashing AdipoRon's Potential: A Fresh Approach to Tackle Pseudomonas aeruginosa Infections in Bronchiectasis via Sphingosine Metabolism Modulation.J Inflamm Res. 2024 Oct 24;17:7653-7674. doi: 10.2147/JIR.S483689. eCollection 2024. J Inflamm Res. 2024. PMID: 39469062 Free PMC article.
-
Small Proline-Rich Protein 3 Regulates IL-33/ILC2 Axis to Promote Allergic Airway Inflammation.Front Immunol. 2022 Jan 20;12:758829. doi: 10.3389/fimmu.2021.758829. eCollection 2021. Front Immunol. 2022. PMID: 35126350 Free PMC article.
-
Environmental allergens house dust mite-induced asthma is associated with ferroptosis in the lungs.Exp Ther Med. 2021 Dec;22(6):1483. doi: 10.3892/etm.2021.10918. Epub 2021 Oct 26. Exp Ther Med. 2021. PMID: 34765024 Free PMC article.
-
Hypermethylation of RNF125 promotes autophagy-induced oxidative stress in asthma by increasing HMGB1 stability.iScience. 2023 Jul 27;26(8):107503. doi: 10.1016/j.isci.2023.107503. eCollection 2023 Aug 18. iScience. 2023. PMID: 37599832 Free PMC article.
-
Intranasal Administration of Recombinant Mycobacterium smegmatis Inducing IL-17A Autoantibody Attenuates Airway Inflammation in a Murine Model of Allergic Asthma.PLoS One. 2016 Mar 14;11(3):e0151581. doi: 10.1371/journal.pone.0151581. eCollection 2016. PLoS One. 2016. PMID: 26974537 Free PMC article.
References
-
- Bochner, B.S., B.J. Undem, and L.M. Lichtenstein. 1994. Immunological aspects of allergic asthma. Annu. Rev. Immunol. 12:295–335. - PubMed
-
- Robinson, D., Q. Hamid, S. Ying, A. Bentley, B. Assoufi, S. Durham, and A.B. Kay. 1993. Prednisolone treatment in asthma is associated with modulation of bronchoalveolar lavage cell interleukin-4, interleukin-5, and interferon-γ cytokine gene expression. Am. Rev. Respir. Dis. 148:401–406. - PubMed
-
- Weiss, J.W., J.M. Drazen, N. Coles, E.R. McFadden, Jr., P.F. Weller, E.J. Corey, R.A. Lewis, and K.F. Austen. 1982. Bronchoconstrictor effects of leukotriene C in humans. Science. 216:196–198. - PubMed
-
- Cuss, F.M., C.M. Dixon, and P.J. Barnes. 1986. Effects of inhaled platelet activating factor on pulmonary function and bronchial responsiveness in man. Lancet. 2:189–192. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
