

Antigenic peptides complexed to phylogenically diverse Hsp70s induce differential immune responses
- PMID: 14627199
- PMCID: PMC514865
- DOI: 10.1379/1466-1268(2003)008<0134:apctpd>2.0.co;2
Antigenic peptides complexed to phylogenically diverse Hsp70s induce differential immune responses
Abstract
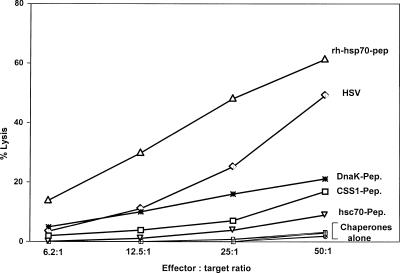
The Hsp70 class of heat shock proteins (Hsps) has been implicated at multiple points in the immune response, including initiation of proinflammatory cytokine production, antigen recognition and processing, and phenotypic maturation of antigen-presenting cells (APCs). This class of chaperones is highly conserved in both sequence and structure, from prokaryotes to higher eukaryotes. In all cases, these chaperones function to bind short segments of either peptides or proteins through an adenosine triphosphate-dependent process. In addition to a possible role in antigen presentation, these chaperones have also been proposed to function as a potent adjuvant. We compared 4 evolutionary diverse Hsp70s, E. coli DnaK, wheat cytosolic Hsc70, plant chloroplastic CCS1, and human Hsp70, for their ability to prime and augment a primary immune response against herpes simplex virus-1 (HSV1). We discovered that all 4 Hsp70s were highly effective as adjuvants displaying similar ability to lipopolysaccharides in upregulating cytokine gene expression. In addition, they were all capable of inducing phenotypic maturation of APCs, as measured by the display of various costimulatory molecules. However, only the human Hsp70 was able to mediate sufficient cross-priming activity to afford a protective immune response to HSV1, as judged by protection from a lethal viral challenge, in vitro proliferation, cytotoxicity, and intracellular interferon-gamma production. The difference in immune response generated by the various Hsp70s could possibly be due to their differential ability to interact productively with other coreceptors and different regulatory cochaperones.
Figures




References
-
- Asea A, Kraeft SK, Kurt-Jones EA, Stevenson MA, Chen LB, Finberg RW, Koo GC, Calderwood SK. HSP70 stimulates cytokine production through a CD14-dependant pathway, demonstrating its dual role as a chaperone and cytokine. Nat Med. 2000;6:435–442. - PubMed
-
- Basu S, Binder RJ, Ramalingam T, Srivastava PK. CD91 is a common receptor for heat shock proteins gp96, hsp90, hsp70, and calreticulin. Immunity. 2001;14:303–313. - PubMed
-
- Basu S, Binder RJ, Suto R, Anderson KM, Srivastava PK. Necrotic but not apoptotic cell death releases heat shock proteins, which deliver a partial maturation signal to dendritic cells and activate the NF-kappa B pathway. Int Immunol. 2000;12:1539–1546. - PubMed
-
- Beech JT, Siew LK, Ghoraishian M, Stasiuk LM, Elson CJ, Thompson SJ. CD4+ Th2 cells specific for mycobacterial 65-kilodalton heat shock protein protect against pristane-induced arthritis. J Immunol. 1997;159:3692–3697. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous