

Perisynaptic localization of delta subunit-containing GABA(A) receptors and their activation by GABA spillover in the mouse dentate gyrus
- PMID: 14627650
- PMCID: PMC6740905
- DOI: 10.1523/JNEUROSCI.23-33-10650.2003
Perisynaptic localization of delta subunit-containing GABA(A) receptors and their activation by GABA spillover in the mouse dentate gyrus
Abstract
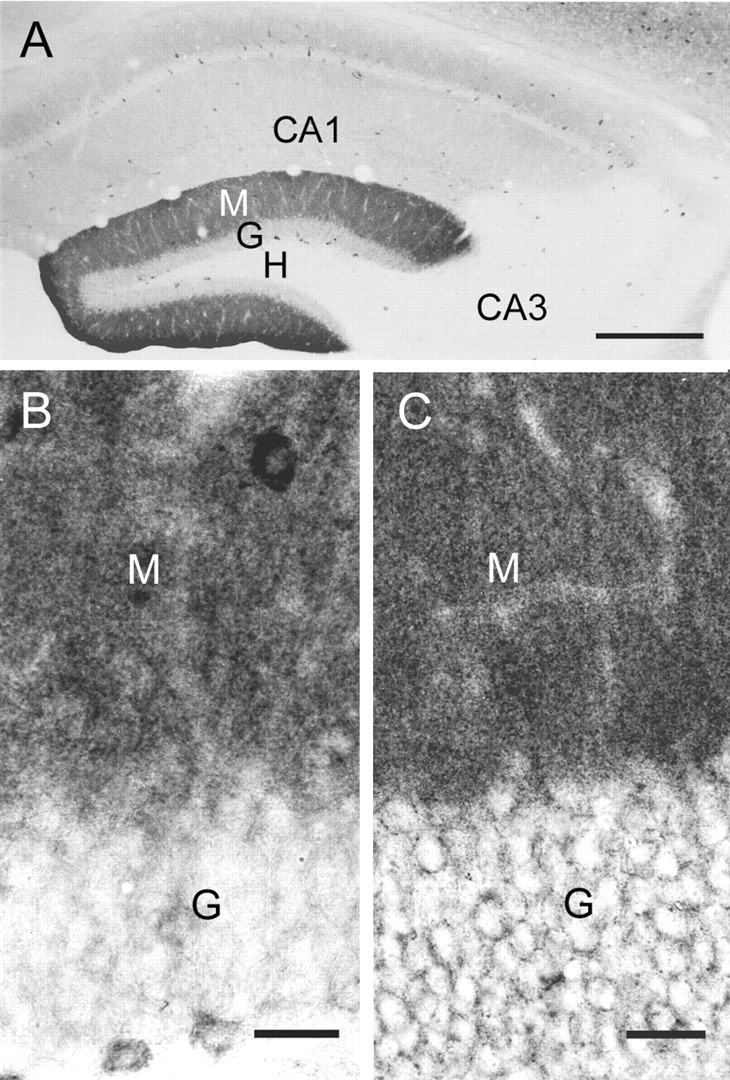
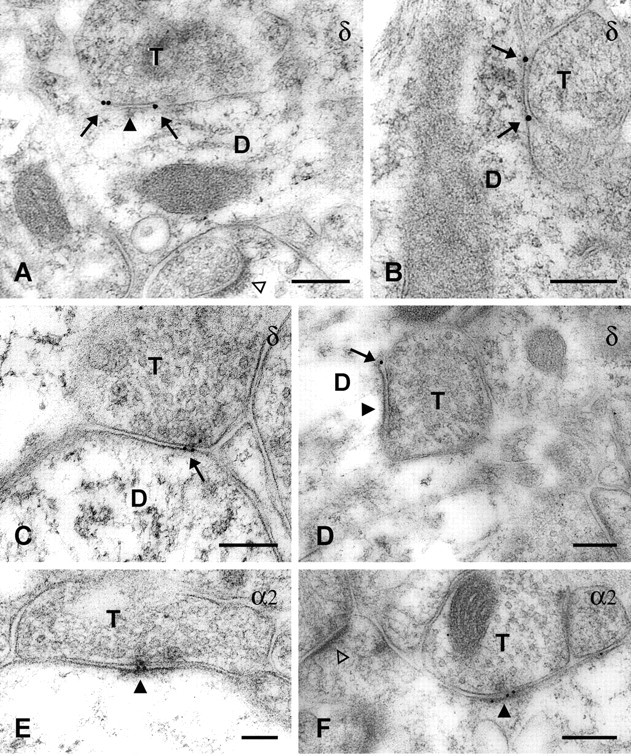
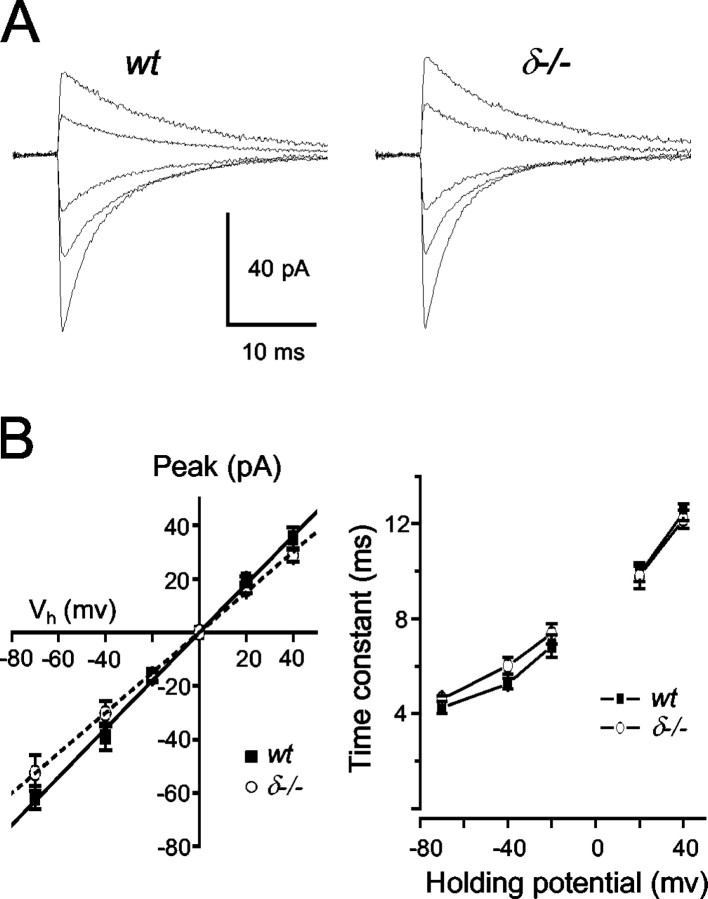
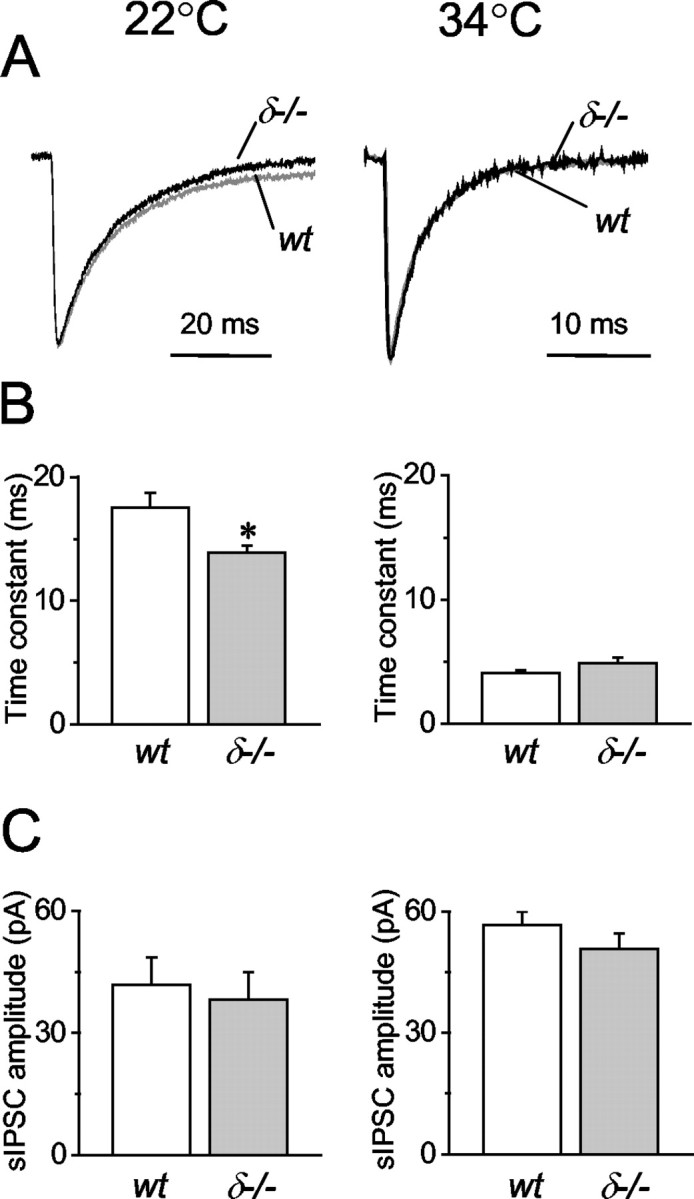
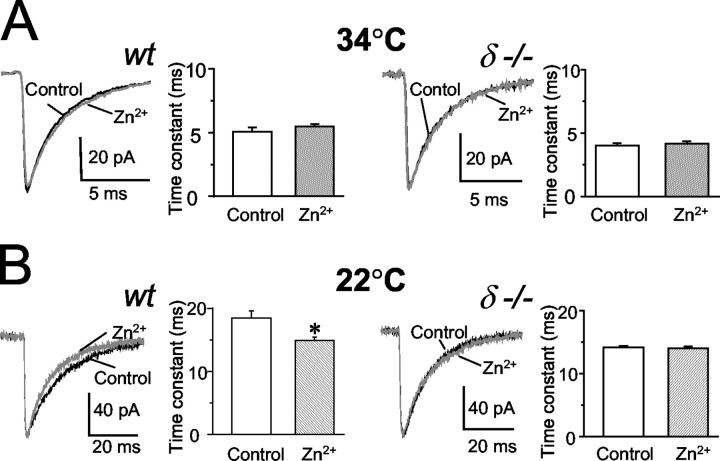
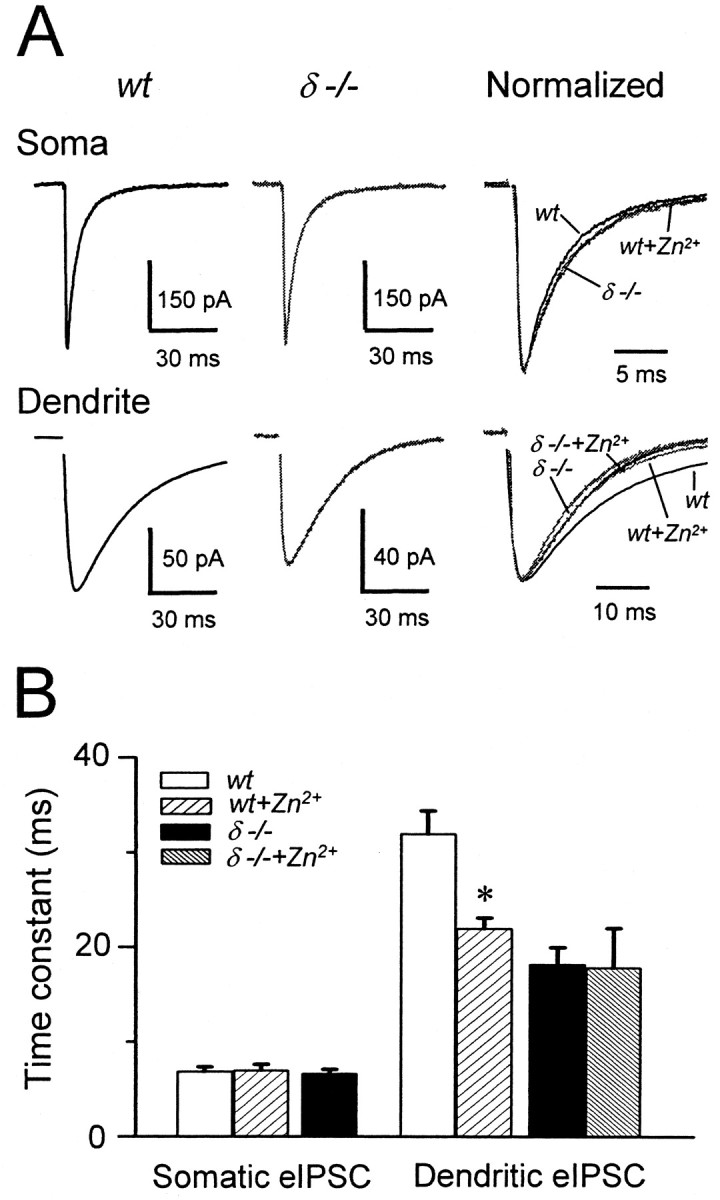
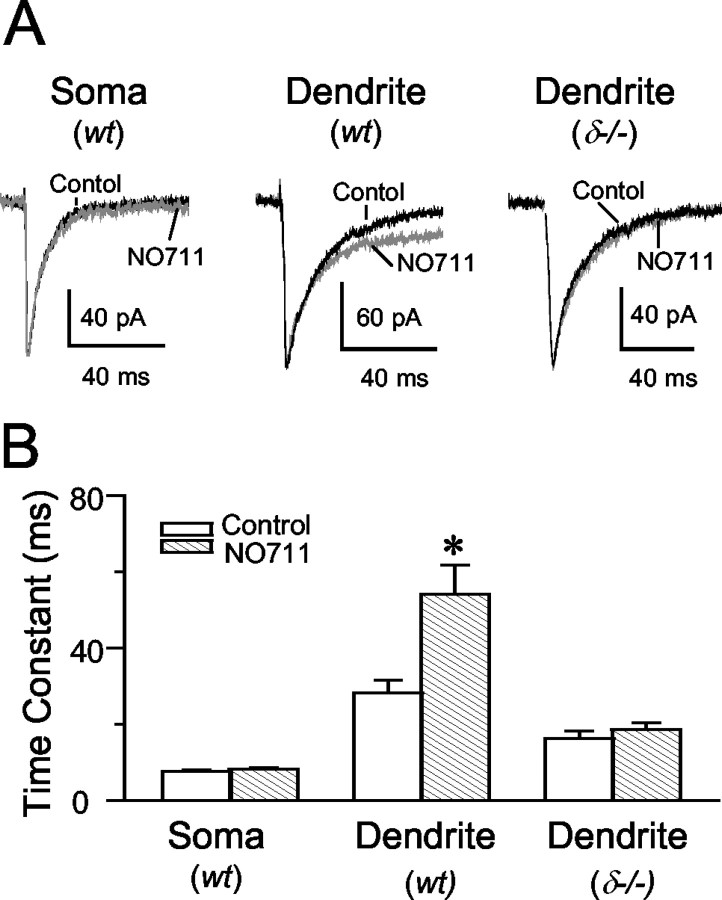
In cerebellar granule cells, delta subunit-containing GABA(A) receptors are found exclusively at extrasynaptic sites, but their subcellular distribution in other brain areas is poorly understood. We examined the anatomical localization and physiological activation of these receptors in adult mouse dentate gyrus granule cells. Immunocytochemistry revealed a high density of delta subunits in the molecular layer and a much lower density in the cell body layer. At the ultrastructural level, immunogold-labeled delta subunits were found at the edge of symmetric synapses on granule cell dendrites. Functional correlates of this perisynaptic localization were obtained by comparing inhibitory responses in delta subunit-deficient (delta-/-) and wild-type (wt) mice. In whole-cell recordings at 22 degrees C, the weighted decay time constants (tau(w)) of spontaneous IPSCs (sIPSCs) were significantly longer in wt mice but were similar at 34 degrees C, reflecting the role of temperature-dependent GABA uptake in shaping sIPSC decay. IPSCs evoked by minimal stimulation (eIPSCs) near the somata had similar tau(w) in delta-/- and wt mice, but eIPSCs elicited from dendritic sites decayed significantly more slowly in wt mice, consistent with a higher density of delta subunit-containing receptors in the molecular layer. The tau(w) of dendritic eIPSCs of wt mice were shortened by ZnCl2 (10 microm), reflecting the high Zn2+ sensitivity of delta subunit-containing GABA(A) receptors, and were prolonged by the GAT-1 GABA transporter inhibitor NO711 (10 microm). Our results demonstrate a perisynaptic localization of delta subunit-containing GABA(A) receptors and indicate that these receptors can be activated by GABA overspill in the molecular layer.
Figures
Similar articles
-
Subtype-specific GABA transporter antagonists synergistically modulate phasic and tonic GABAA conductances in rat neocortex.J Neurophysiol. 2005 Sep;94(3):2073-85. doi: 10.1152/jn.00520.2005. Epub 2005 Jun 29. J Neurophysiol. 2005. PMID: 15987761
-
GABA transporters regulate inhibition in the retina by limiting GABA(C) receptor activation.J Neurosci. 2002 Apr 15;22(8):3285-92. doi: 10.1523/JNEUROSCI.22-08-03285.2002. J Neurosci. 2002. PMID: 11943830 Free PMC article.
-
Altered localization of GABA(A) receptor subunits on dentate granule cell dendrites influences tonic and phasic inhibition in a mouse model of epilepsy.J Neurosci. 2007 Jul 11;27(28):7520-31. doi: 10.1523/JNEUROSCI.1555-07.2007. J Neurosci. 2007. PMID: 17626213 Free PMC article.
-
Regulation of GABAA receptor trafficking, channel activity, and functional plasticity of inhibitory synapses.Pharmacol Ther. 2004 Jun;102(3):195-221. doi: 10.1016/j.pharmthera.2004.04.003. Pharmacol Ther. 2004. PMID: 15246246 Review.
-
Function and modulation of delta-containing GABA(A) receptors.Psychoneuroendocrinology. 2009 Dec;34 Suppl 1:S67-73. doi: 10.1016/j.psyneuen.2009.08.010. Psychoneuroendocrinology. 2009. PMID: 19766404 Free PMC article. Review.
Cited by
-
The population firing rate in the presence of GABAergic tonic inhibition in single neurons and application to general anaesthesia.Cogn Neurodyn. 2012 Jun;6(3):227-37. doi: 10.1007/s11571-011-9182-9. Epub 2011 Nov 17. Cogn Neurodyn. 2012. PMID: 23730354 Free PMC article.
-
γ-Aminobutyric acid type A (GABAA) receptor α subunits play a direct role in synaptic versus extrasynaptic targeting.J Biol Chem. 2012 Aug 10;287(33):27417-30. doi: 10.1074/jbc.M112.360461. Epub 2012 Jun 18. J Biol Chem. 2012. PMID: 22711532 Free PMC article.
-
Normal and epilepsy-associated pathologic function of the dentate gyrus.Prog Brain Res. 2016;226:155-78. doi: 10.1016/bs.pbr.2016.04.005. Epub 2016 May 18. Prog Brain Res. 2016. PMID: 27323942 Free PMC article. Review.
-
Novel types of frequency filtering in the lateral perforant path projections to dentate gyrus.J Physiol. 2022 Aug;600(16):3865-3896. doi: 10.1113/JP283012. Epub 2022 Aug 1. J Physiol. 2022. PMID: 35852108 Free PMC article.
-
Single Ethanol Withdrawal Regulates Extrasynaptic δ-GABAA Receptors Via PKCδ Activation.Front Mol Neurosci. 2018 Apr 26;11:141. doi: 10.3389/fnmol.2018.00141. eCollection 2018. Front Mol Neurosci. 2018. PMID: 29755316 Free PMC article.
References
-
- Borden LA, Murali Dhar TG, Smith KE, Weinshank RL, Branchek TA, Gluchowski C ( 1994) Tiagabine, SK&F 89976-A, CI-966, and NNC-711 are selective for the cloned GABA transporter GAT-1. Eur J Pharmacol 269: 219-224. - PubMed
-
- Brickley SG, Revilla V, Cull-Candy SG, Wisden W, Farrant M ( 2001) Adaptive regulation of neuronal excitability by a voltage-independent potassium conductance. Nature 409: 88-92. - PubMed
-
- Brooks-Kayal AR, Shumate MD, Jin H, Rikhter TY, Coulter DA ( 1998) Selective changes in single cell GABAA receptor subunit expression and function in temporal lobe epilepsy. Nat Med 4: 1166-1172. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases