

Comparative Study
doi: 10.1101/gad.1140303.
Template boundary definition in mammalian telomerase
Affiliations
- PMID: 14630939
- PMCID: PMC280623
- DOI: 10.1101/gad.1140303
Item in Clipboard
Comparative Study
Template boundary definition in mammalian telomerase
Genes Dev.
.
Abstract
Telomerase uses a short template sequence in its intrinsic RNA component to synthesize telomere repeats. Disruption of the helix P1b in human telomerase RNA or alteration of its distance from the template resulted in telomerase copying residues past the normal template boundary both in vivo and in vitro. Therefore, helix P1b is important for template boundary definition in human telomerase. Mouse telomerase RNA lacks helix P1b, and the boundary is established at 2 nt downstream of the 5'-end. The divergent structure of boundary definition elements in mammals, yeast, and ciliates suggests diverse mechanisms for template boundary definition in telomerase.
Figures

Effects of 5′-end truncation of hTR on telomerase activity. (A) Schematic diagram of the secondary structure of hTR. The nucleotide positions of the 5′-end (1, 15, 26, 32, 44) and the 3′-end (212) of truncated RNA mutants are indicated. Each helical region is labeled as previously described (Chen et al. 2000, 2002). The RNA fragments, pseudoknot and CR4–CR5 fragments, used in the in vitro reconstitution of telomerase are indicated by a dashed bracket and a dashed box, respectively. The conserved pseudoknot structural domain is also indicated by a dashed box. (B) Telomerase activity assay of telomerase reconstituted with the 5′-truncated telomerase RNA fragments. The truncated RNA fragments, as indicated above each lane, were reconstituted with in vitro expressed hTERT-HA2 protein in the presence of the CR4–CR5 RNA fragment. Immunopurified enzyme was assayed for telomerase activity using telomere primer (TTAGGG)3 in the presence of all four nucleotides (dNTPs). For each major band, the number of nucleotides added to the 3′-end of the primer is indicated either on the left side (+4, +10, +16, +22) or on the right side (+6, +12, +18, +24) of the gel.
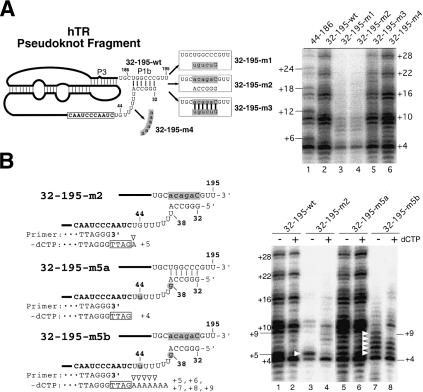
Effects of mutations in the P1b helix of hTR on telomerase template boundary definition. (A, left) Schematic diagram of the mutant hTR pseudoknot fragments. The regions altered in each pseudoknot mutant are shaded, and the sequences changed are shown in lowercase. (Right) Telomerase activity assay with in vitro reconstituted telomerase containing the P1b mutant RNAs. Each pseudoknot RNA fragment with a P1b mutation, as indicated above each lane, was reconstituted with hTERT-HA2 protein and an hTR CR4–CR5 RNA fragment. Immunopurified enzyme was assayed for telomerase activity in the presence of all four nucleotides (dNTPs). (B, left) Schematic diagram of the mutant hTR pseudoknot fragments. The regions altered in each pseudoknot mutant are shaded, and the sequences changed are shown in lowercase. The primer sequence is aligned with the RNA template sequence that is shown in bold. Nucleotides in the boxes represent nucleotides added to the 3′-end of primers. Open triangles indicate nucleotides added with nontelomeric sequence. (Right) Telomerase activity assay of in vitro reconstituted telomerase containing mutant telomerase RNA fragments. Reconstituted enzyme was assayed for telomerase activity in either the absence or the presence of 0.5 mM dCTP. White triangles indicate bands of read-through product with nontelomeric nucleotides incorporated.
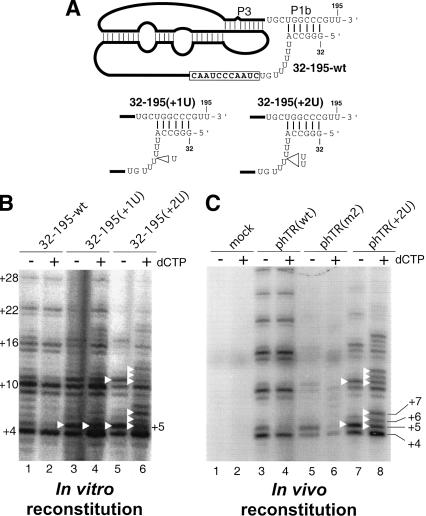
Effects of altered linker length between the P1b helix and the template on template boundary definition. (A) Schematic diagram of the mutant hTR pseudoknot fragments. The U-residue insertions made in the pseudoknot mutants, 32–195(+1U) and 32–195(+2U), are indicated by open triangles. (B) Telomerase activity assay of in vitro reconstituted telomerase containing the linker-length mutant RNAs. Various pseudoknot RNA fragments— hTR32–195-wt, hTR32–195(+1U), and hTR32–195(+2U)—were assembled with hTR–CR4–CR5 RNA and in vitro expressed hTERTHA protein. In vitro reconstituted enzyme was immunopurified and assayed for activity. The activity assay was carried out in either the absence or the presence of 0.5 mM dCTP. Bands representing incorporation past the normal template are marked with white triangles. (C) Telomerase activity assay of in vivo reconstituted human mutant telomerase. The telomerase RNA genes are expressed in vivo from phTR(wt), phTR(m2), or phTR(+2U), as indicated above the gel. A mock transfection with only phTERT-HA was included to ensure the absence of endogenous hTR expression (lanes 1,2).

Effects of 5′-end extension of mouse telomerase RNA on telomerase template boundary definition. (A) Analysis of in vitro reconstituted mouse telomerase. (Top) Schematic diagram of the mutant mTR pseudoknot fragments used in the in vitro reconstitution. The sequence changed at the 5′-end of each RNA mutant— mTR(+1g)-155, mTR(-2g)-155, mTR(-4g)-155, and mTR(-13)-155(P1)—is shaded. (Bottom) Direct activity assay of in vitro reconstituted mouse telomerase. Each pseudoknot RNA fragment with either a 5′-end extension or the artificial P1b helix was reconstituted with the mTERT-HA2 protein and the mTR CR4–CR5 RNA fragment, and assayed for activity in the presence or absence of dCTP. (B) Analysis of in vivo reconstituted mouse telomerase. (Top) Sequence at the 5′-end of the mTR mutants. The sequence changed at the 5′-end of each RNA mutant—mTR(+1g) and mTR(add2+1g)—is shaded. The template boundary located 2 nt downstream of the 5′-end is indicated with an arrow. The primer sequence is aligned with the RNA template sequence that is shown in bold. Nucleotides in the box represent nucleotides added at the 3′-end of primers. Open triangles indicate nucleotides added past the normal boundary. (Bottom) Direct activity assay of in vivo reconstituted mouse telomerase with the mTR mutant. Mouse telomerase RNAs were expressed from pmTR(wt), pmTR(+1g), and pmTR(add2+1g), and reconstituted with mTERT-HA protein in mTR-deficient mouse embryonic fibroblast cells. The reconstituted enzyme was immunopurified and assayed for telomerase assay in the presence or absence of dCTP. The number of nucleotides added at the major bands is marked on the right of the gel.

(A) Comparison of template boundary definition elements among human, mouse, yeast, and ciliate telomerase RNAs. The known secondary structures of regions flanking the RNA template are depicted for human, mouse, Kluyveromyces lactis (yeast), and Tetrahymena thermophila (ciliate) telomerase RNAs. The sequence of the template region is shown in a black box, and the RNA structure essential for template boundary definition is shown in a red box with a red line to indicate function in boundary definition. (B) A proposed model for template boundary definition in human telomerase. The TERT protein is depicted as a gray sphere with the catalytic site shown as an open circle. The helix P1b (shown in blue) serves as an anchor point for tethering the linker (shown in blue) and the template region (white box). During the copying of the template, the movement of the RNA template within the catalytic site is constrained by the length of the linker region.
References
-
- Autexier C. and Greider, C.W. 1995. Boundary elements of the Tetrahymena telomerase RNA template and alignment domains. Genes & Dev. 9: 2227-2239. - PubMed
-
- Avilion A.A. 1995. Characterization and expression of human telomerase. In Graduate Program in Cellular and Developmental Biology, pp. 234. State University of New York at Stony Brook, Stony Brook, NY.
-
- Beattie T.L., Zhou, W., Robinson, M.O., and Harrington, L. 1998. Reconstitution of human telomerase activity in vitro. Curr. Biol. 8: 177-180. - PubMed
-
- Blasco M.A., Lee, H.W., Hande, M.P., Samper, E., Lansdorp, P.M., DePinho, R.A., and Greider, C.W. 1997. Telomere shortening and tumor formation by mouse cells lacking telomerase RNA. Cell 91: 25-34. - PubMed
-
- Bryan T.M., Goodrich, K.J., and Cech, T.R. 2000. Telomerase RNA bound by protein motifs specific to telomerase reverse transcriptase. Mol. Cell 6: 493-499. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous