

Lipoprotein receptors and a disabled family cytoplasmic adaptor protein regulate EGL-17/FGF export in C. elegans
- PMID: 14630941
- PMCID: PMC280628
- DOI: 10.1101/gad.1136103
Lipoprotein receptors and a disabled family cytoplasmic adaptor protein regulate EGL-17/FGF export in C. elegans
Abstract
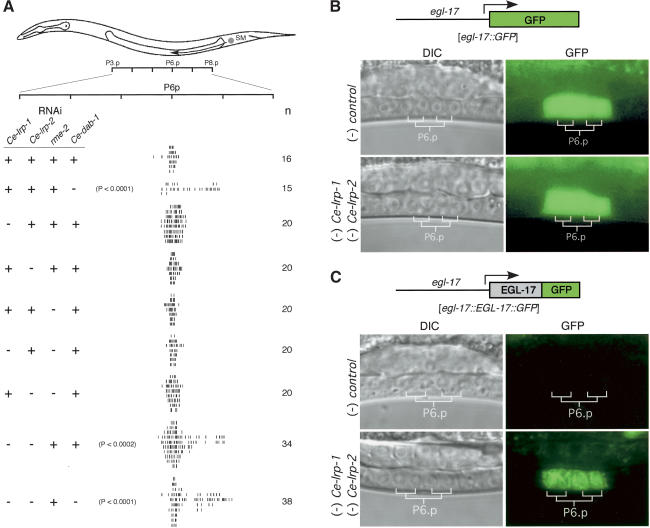
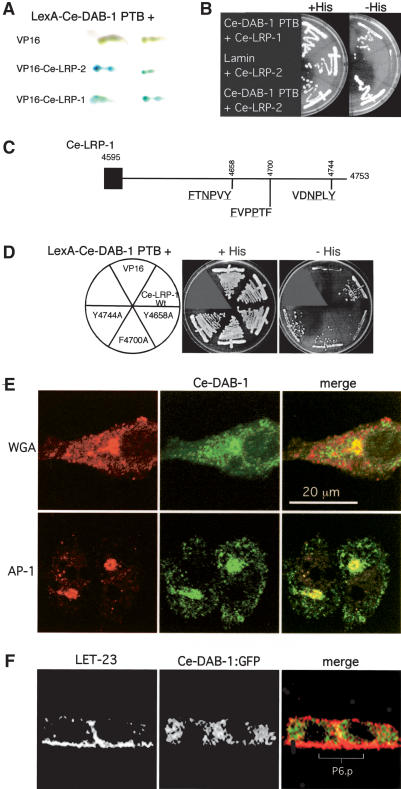
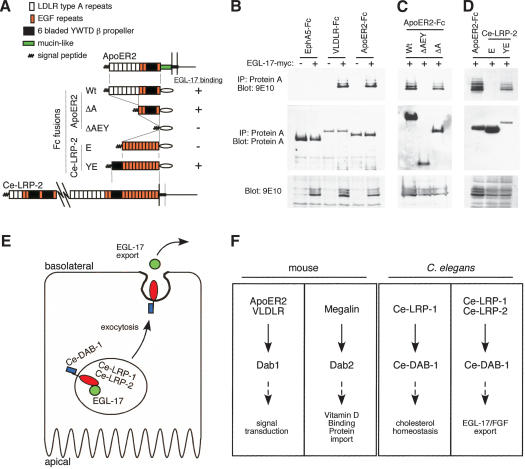
Growth factors and morphogens need to be secreted to act on distant cells during development and in response to injury. Here, we report evidence that efficient export of a fibroblast growth factor (FGF), EGL-17, from the Caenorhabditis elegans developing vulva requires the lipoprotein receptor-related proteins Ce-LRP-1 and Ce-LRP-2 and a cytoplasmic adaptor protein, Ce-DAB-1 (Disabled). Lipoprotein receptors are transmembrane proteins best known for their roles in endocytosis. Ce-LRP-1 and Ce-LRP-2 possess a conserved intraluminal domain that can bind to EGL-17, as well as a cytosolic FXNPXY motif that can bind to Ce-DAB-1. Ce-DAB-1 contains signals that confer subcellular localization to Golgi-proximal vesicles. These results suggest a model in which Ce-DAB-1 coordinates selection of receptors and cargo, including EGL-17, for transport through the secretory pathway.
Figures



Similar articles
-
Clathrin interaction and subcellular localization of Ce-DAB-1, an adaptor for protein secretion in Caenorhabditis elegans.Traffic. 2006 Mar;7(3):324-36. doi: 10.1111/j.1600-0854.2006.00386.x. Traffic. 2006. PMID: 16497226
-
EGL-17(FGF) expression coordinates the attraction of the migrating sex myoblasts with vulval induction in C. elegans.Development. 1998 Mar;125(6):1083-93. doi: 10.1242/dev.125.6.1083. Development. 1998. PMID: 9463355
-
Cell fates and fusion in the C. elegans vulval primordium are regulated by the EGL-18 and ELT-6 GATA factors -- apparent direct targets of the LIN-39 Hox protein.Development. 2002 Nov;129(22):5171-80. doi: 10.1242/dev.129.22.5171. Development. 2002. PMID: 12399309
-
A pair as a minimum: the two fibroblast growth factors of the nematode Caenorhabditis elegans.Dev Dyn. 2005 Feb;232(2):247-55. doi: 10.1002/dvdy.20219. Dev Dyn. 2005. PMID: 15614779 Review.
-
Control of cell migration during Caenorhabditis elegans development.Curr Opin Cell Biol. 1999 Oct;11(5):608-13. doi: 10.1016/s0955-0674(99)00028-9. Curr Opin Cell Biol. 1999. PMID: 10508660 Review.
Cited by
-
Repulsive guidance molecule acts in axon branching in Caenorhabditis elegans.Sci Rep. 2021 Nov 16;11(1):22370. doi: 10.1038/s41598-021-01853-8. Sci Rep. 2021. PMID: 34785759 Free PMC article.
-
Low-Density Lipoprotein Receptor LRP-2 regulates GLR-1 glutamate receptors and glutamatergic behavior in C. elegans.MicroPubl Biol. 2023 Apr 26;2023:10.17912/micropub.biology.000837. doi: 10.17912/micropub.biology.000837. eCollection 2023. MicroPubl Biol. 2023. PMID: 37179968 Free PMC article.
-
Regulated trafficking of the MSP/Eph receptor during oocyte meiotic maturation in C. elegans.Curr Biol. 2008 May 20;18(10):705-714. doi: 10.1016/j.cub.2008.04.043. Epub 2008 May 8. Curr Biol. 2008. PMID: 18472420 Free PMC article.
-
Canonical RTK-Ras-ERK signaling and related alternative pathways.WormBook. 2013 Jul 11:1-38. doi: 10.1895/wormbook.1.80.2. WormBook. 2013. PMID: 23908058 Free PMC article. Review.
-
Cargo- and compartment-selective endocytic scaffold proteins.Biochem J. 2004 Oct 1;383(Pt 1):1-11. doi: 10.1042/BJ20040913. Biochem J. 2004. PMID: 15219178 Free PMC article. Review.
References
-
- Amanai K. and Jiang, J. 2001. Distinct roles of Central missing and Dispatched in sending the Hedgehog signal. Development 128: 5119-5127. - PubMed
-
- Borland C.Z., Schutzman, J.L., and Stern, M.J. 2001. Fibroblast growth factor signaling in Caenorhabditis elegans. BioEssays 23: 1120-1130. - PubMed
-
- Branda C.S. and Stern, M.J. 2000. Mechanisms controlling sex myoblast migration in Caenorhabditis elegans hermaphrodites. Dev. Biol. 226: 137-151. - PubMed
-
- Burdine R.D., Branda, C.S., and Stern, M.J. 1998. EGL-17(FGF) expression coordinates the attraction ofthe migrating sex myoblasts with vulval induction in C. elegans. Development 125: 1083-1093. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous