

Photosynthesis and state transitions in mitochondrial mutants of Chlamydomonas reinhardtii affected in respiration
- PMID: 14630958
- PMCID: PMC300752
- DOI: 10.1104/pp.103.028076
Photosynthesis and state transitions in mitochondrial mutants of Chlamydomonas reinhardtii affected in respiration
Abstract
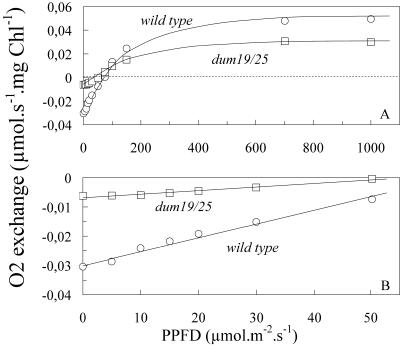
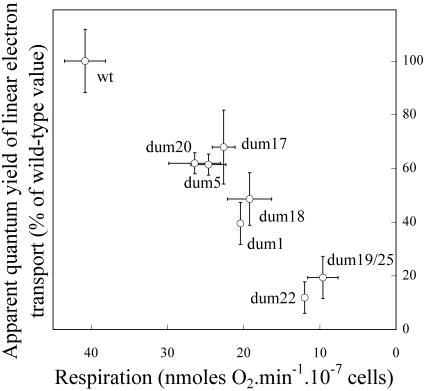
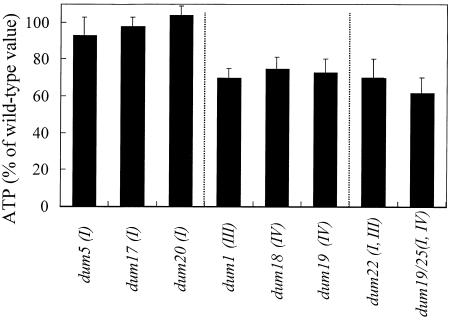
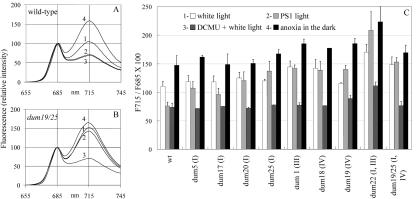
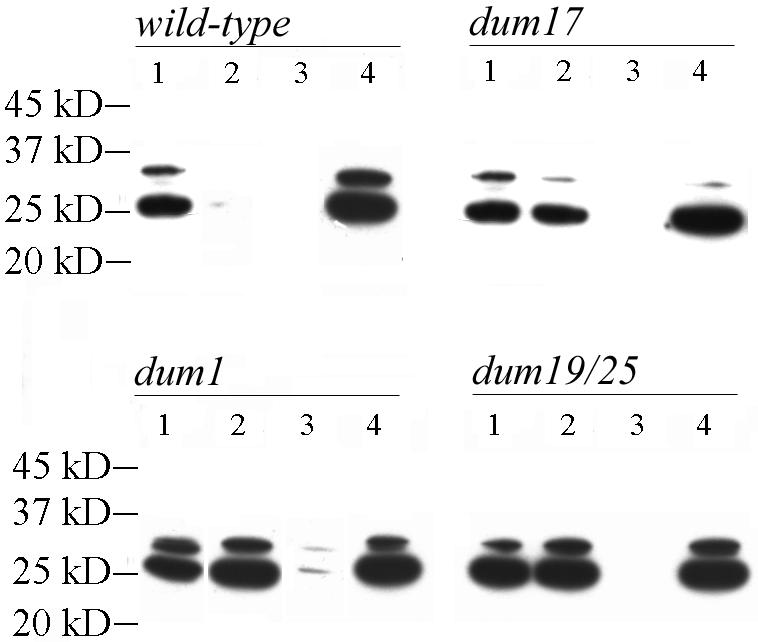
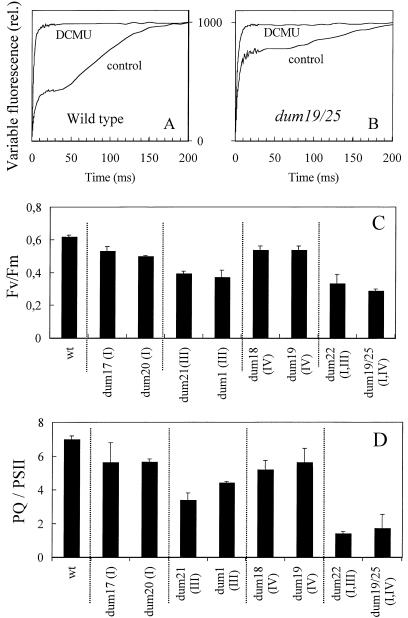
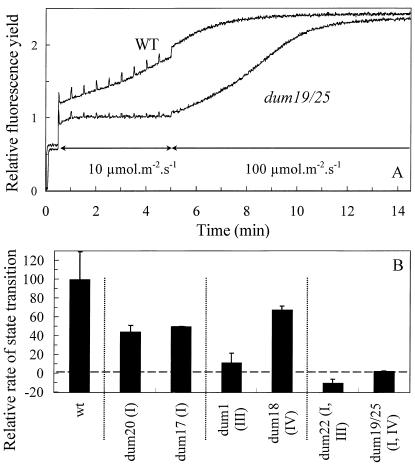
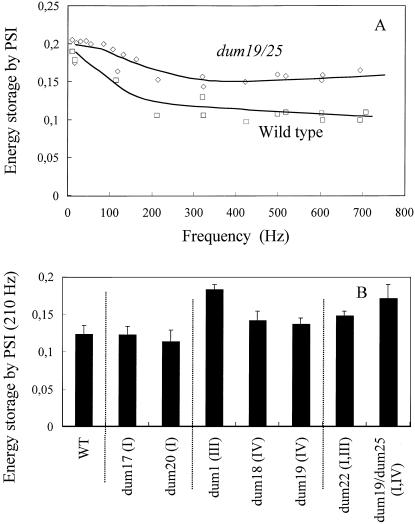
Photosynthetic activities were analyzed in Chlamydomonas reinhardtii mitochondrial mutants affected in different complexes (I, III, IV, I + III, and I + IV) of the respiratory chain. Oxygen evolution curves showed a positive relationship between the apparent yield of photosynthetic linear electron transport and the number of active proton-pumping sites in mitochondria. Although no significant alterations of the quantitative relationships between major photosynthetic complexes were found in the mutants, 77 K fluorescence spectra showed a preferential excitation of photosystem I (PSI) compared with wild type, which was indicative of a shift toward state 2. This effect was correlated with high levels of phosphorylation of light-harvesting complex II polypeptides, indicating the preferential association of light-harvesting complex II with PSI. The transition to state 1 occurred in untreated wild-type cells exposed to PSI light or in 3-(3,4-dichlorophenyl)-1,1-dimethylureatreated cells exposed to white light. In mutants of the cytochrome pathway and in double mutants, this transition was only observed in white light in the presence of 3-(3,4-dichlorophenyl)-1,1-dimethylurea. This suggests higher rates of nonphotochemical plastoquinone reduction through the chlororespiratory pathway, which was confirmed by measurements of the complementary area above the fluorescence induction curve in dark-adapted cells. Photo-acoustic measurements of energy storage by PSI showed a stimulation of PSI-driven cyclic electron flow in the most affected mutants. The present results demonstrate that in C. reinhardtii mutants, permanent defects in the mitochondrial electron transport chain stabilize state 2, which favors cyclic over linear electron transport in the chloroplast.
Figures
Similar articles
-
Excitation energy transfer in Chlamydomonas reinhardtii deficient in the PSI core or the PSII core under conditions mimicking state transitions.Biochim Biophys Acta. 2016 Jun;1857(6):625-33. doi: 10.1016/j.bbabio.2016.03.002. Epub 2016 Mar 3. Biochim Biophys Acta. 2016. PMID: 26946087
-
Photosynthetic membrane organization and role of state transition in cyt, cpII, stt7 and npq mutants of Chlamydomonas reinhardtii.J Photochem Photobiol B. 2014 Aug;137:77-83. doi: 10.1016/j.jphotobiol.2014.03.025. Epub 2014 Apr 24. J Photochem Photobiol B. 2014. PMID: 24836759
-
Strategies to Study Dark Growth Deficient or Slower Mutants in Chlamydomonas reinhardtii.Methods Mol Biol. 2021;2297:125-140. doi: 10.1007/978-1-0716-1370-2_13. Methods Mol Biol. 2021. PMID: 33656676
-
Respiratory-deficient mutants of the unicellular green alga Chlamydomonas: a review.Biochimie. 2014 May;100:207-18. doi: 10.1016/j.biochi.2013.10.006. Epub 2013 Oct 15. Biochimie. 2014. PMID: 24139906 Review.
-
The central role of the green alga Chlamydomonas reinhardtii in revealing the mechanism of state transitions.J Exp Bot. 2005 Jan;56(411):383-8. doi: 10.1093/jxb/erh230. Epub 2004 Aug 27. J Exp Bot. 2005. PMID: 15333639 Review.
Cited by
-
Cyclic electron flow around photosystem I in unicellular green algae.Photosynth Res. 2010 Nov;106(1-2):47-56. doi: 10.1007/s11120-010-9566-4. Epub 2010 Jun 8. Photosynth Res. 2010. PMID: 20532629
-
Characterization of transformed Arabidopsis with altered alternative oxidase levels and analysis of effects on reactive oxygen species in tissue.Plant Physiol. 2005 Dec;139(4):1806-20. doi: 10.1104/pp.105.070763. Epub 2005 Nov 18. Plant Physiol. 2005. PMID: 16299171 Free PMC article.
-
Mitochondria Affect Photosynthetic Electron Transport and Photosensitivity in a Green Alga.Plant Physiol. 2018 Mar;176(3):2305-2314. doi: 10.1104/pp.17.01249. Epub 2017 Dec 28. Plant Physiol. 2018. PMID: 29284743 Free PMC article.
-
Functional rearrangement of the light-harvesting antenna upon state transitions in a green alga.Biophys J. 2015 Jan 20;108(2):261-71. doi: 10.1016/j.bpj.2014.11.3470. Biophys J. 2015. PMID: 25606675 Free PMC article.
-
The lifetime of the oxygen-evolving complex subunit PSBO depends on light intensity and carbon availability in Chlamydomonas.Plant Cell Environ. 2023 Feb;46(2):422-439. doi: 10.1111/pce.14481. Epub 2022 Nov 17. Plant Cell Environ. 2023. PMID: 36320098 Free PMC article.
References
-
- Allen JF (1992) Protein phosphorylation in regulation of photosynthesis. Biochim Biophys Acta 1098: 275-335 - PubMed
-
- Bennoun P (2001) Chlororespiration and the process of carotenoid biosynthesis. Biochim Biophys Acta 1506: 133-142 - PubMed
-
- Bradford MM (1976) A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
