

B-cell antigen-receptor signalling in lymphocyte development
- PMID: 14632637
- PMCID: PMC1783068
- DOI: 10.1111/j.1365-2567.2003.01756.x
B-cell antigen-receptor signalling in lymphocyte development
Abstract
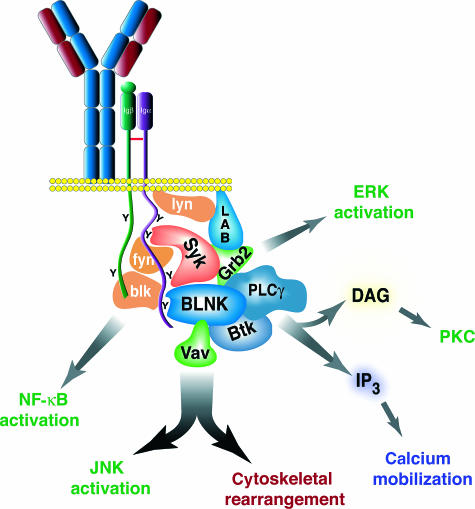
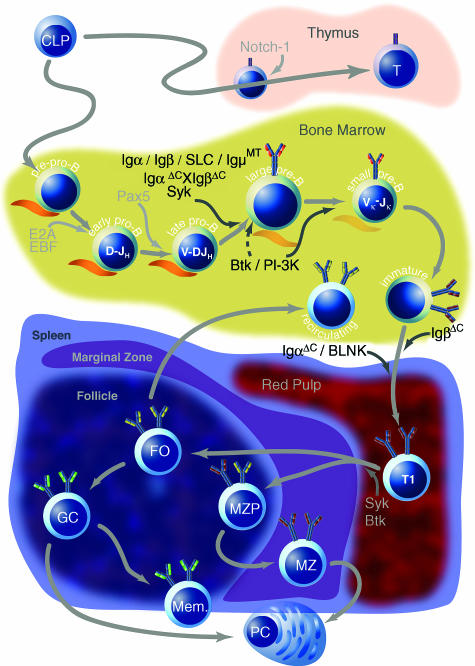
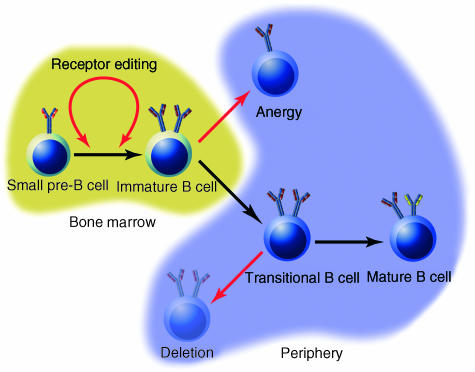
Signalling through the B-cell antigen receptor (BCR) is required throughout B-cell development and peripheral maturation. Targeted disruption of BCR components or downstream effectors indicates that specific signalling mechanisms are preferentially required for central B-cell development, peripheral maturation and repertoire selection. Additionally, the avidity and the context in which antigen is encountered determine both cell fate and differentiation in the periphery. Although the signalling and receptor components required at each stage have been largely elucidated, the molecular mechanisms through which specific signalling are evoked at each stage are still obscure. In particular, it is not known how the pre-BCR initiates the signals required for normal development or how immature B cells regulate the signalling pathways that determine cell fate. In this review, we will summarize the recent studies that have defined the molecules required for B-cell development and maturation as well as the theories on how signals may be regulated at each stage.
Figures
References
-
- Raff MC, Steinberg J, Taylor R. Immunoglobulin determinants on the surface of mouse lymphoid cells. Nature. 1970;225:553. - PubMed
-
- Grupp SA, Campbell K, Mitchell RN, Cambier JC, Abbas AK. Signaling-defective mutants of the B lymphocyte antigen receptor fail to associate with Ig-alpha and Ig-beta/gamma. J Biol Chem. 1993;268:25776. - PubMed
-
- Mitchell RN, Shaw AC, Weaver YK, Leder P, Abbas AK. Cytoplasmic tail deletion converts membrane immunoglobulin to a phosphatidylinositol-linked form lacking signaling and efficient antigen internalization functions. J Biol Chem. 1991;266:8856. - PubMed
-
- Blum JH, Stevens TL, Defranco AL. Role of the mu immunoglobulin heavy chain transmembrane and cytoplasmic domains in B-cell antigen receptor expression and signal transduction. J Biol Chem. 1993;268:27236. - PubMed
-
- Martin SW, Goodnow CC. Burst-enhancing role of the IgG membrane tail as a molecular determinant of memory. Nat Immunol. 2002;3:182. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
