

In vivo expression of interleukin-8, and regulated on activation, normal, T-cell expressed, and secreted, by human germinal centre B lymphocytes
- PMID: 14632656
- PMCID: PMC1783051
- DOI: 10.1046/j.1365-2567.2003.01745.x
In vivo expression of interleukin-8, and regulated on activation, normal, T-cell expressed, and secreted, by human germinal centre B lymphocytes
Abstract
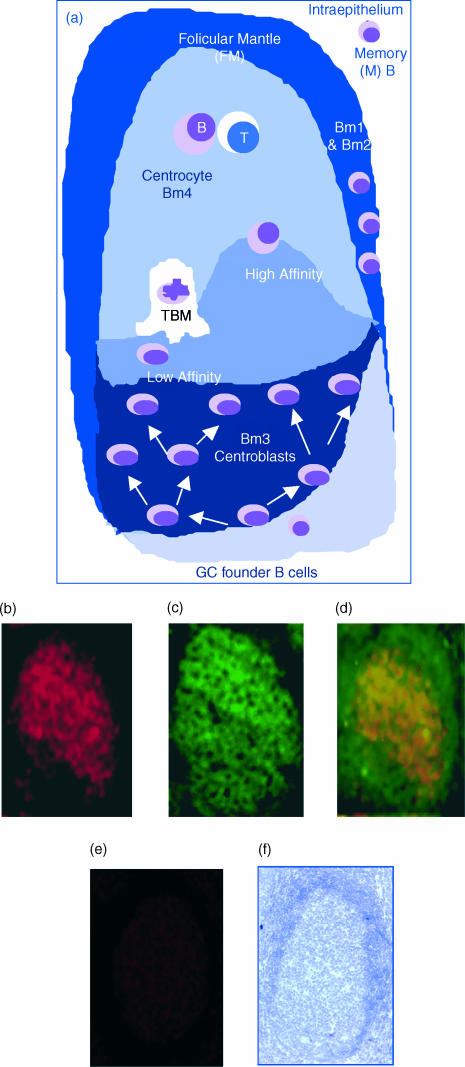
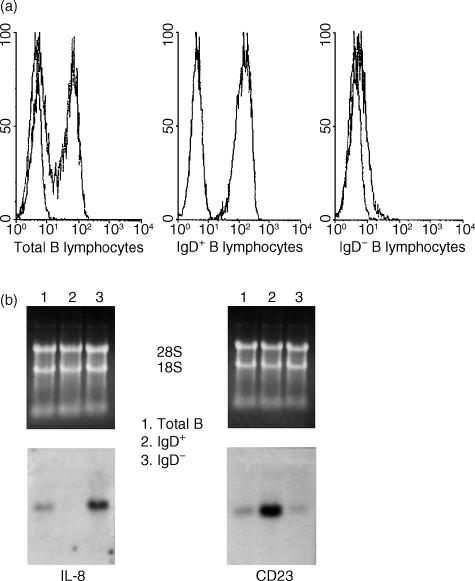
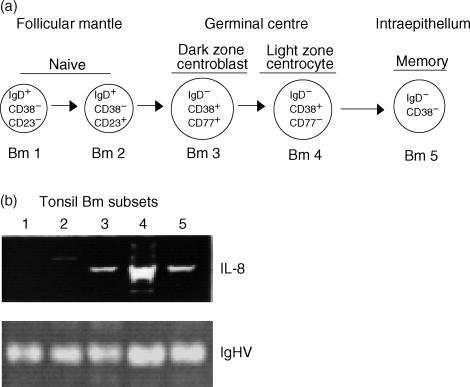
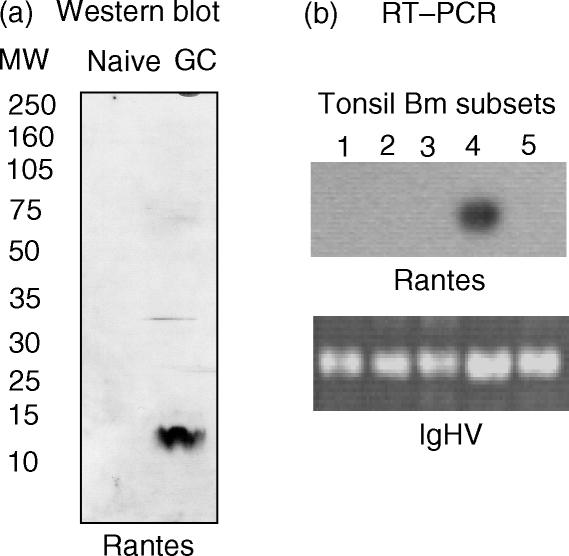
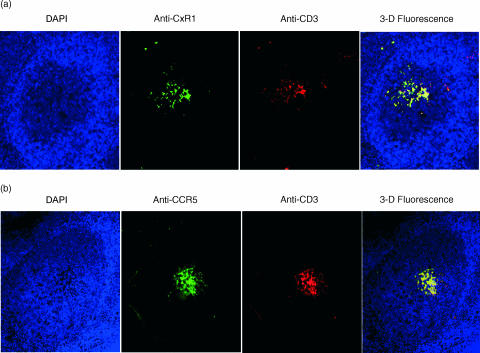
T-cell homing within germinal centres (GCs) is required for humoral B-cell responses. However, the mechanisms implicated in the recruitment of T cells into the GC are not completely understood. Here we show, by immunohistology, and Northern and Western blots, that in vivo human GC B lymphocytes can express CxC and CC chemokines. Moreover, B-cell subset-specific experiments reveal that interleukin (IL)-8 and regulated on activation, normal, T-cell expressed, and secreted (RANTES) are predominantly expressed by GC centroblast and centrocytes, suggesting that chemokine expression is essential at stages in which B-lymphocytes engage in active antigen-dependent interactions with T lymphocytes. In keeping with this hypothesis, we show that the T cells recruited into the GC correlatively express the receptors for IL-8 and RANTES. We propose that chemokine expression is a key B-cell function that facilitates T-lymphocyte recruitment into the GCs and supports cognate B-cell : T-cell encounters. Moreover, our data are consistent with the impaired homing of T cells to secondary lymphoid organs in mice that are either deficient in CC and CxC chemokines or their receptors.
Figures
References
-
- MacLennan ICM. Germinal centers. Annu Rev Immunol. 1994;12:117–39. - PubMed
-
- Kelsoe G. The germinal center: a crucible for lymphocyte selection. Semin Immunol. 1996;8:179–84. - PubMed
-
- Liu Y-J, Zhang J, Lane PJL, Chan EY-T, MacLennan ICM. Sites of specific B-cell activation in primary and secondary responses to T-cell-dependent and T-cell-independent antigens. Eur J Immunol. 1991;21:2951–62. - PubMed
-
- Berek C, Berger A, Apel M. Maturation of immune responses in germinal centers. Cell. 1991;67:1121–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
