

Structural and biochemical analyses of DNA and RNA binding by a bifunctional homing endonuclease and group I intron splicing factor
- PMID: 14633971
- PMCID: PMC289148
- DOI: 10.1101/gad.1109003
Structural and biochemical analyses of DNA and RNA binding by a bifunctional homing endonuclease and group I intron splicing factor
Abstract
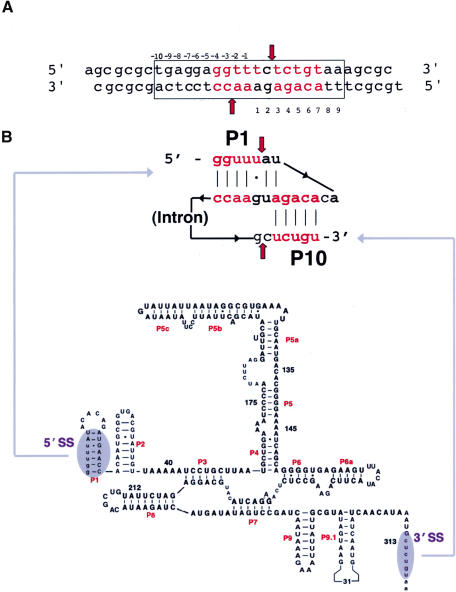
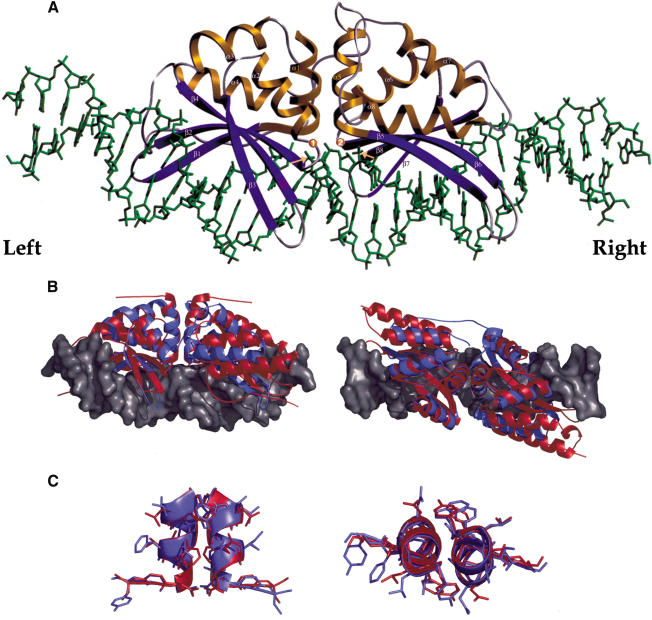
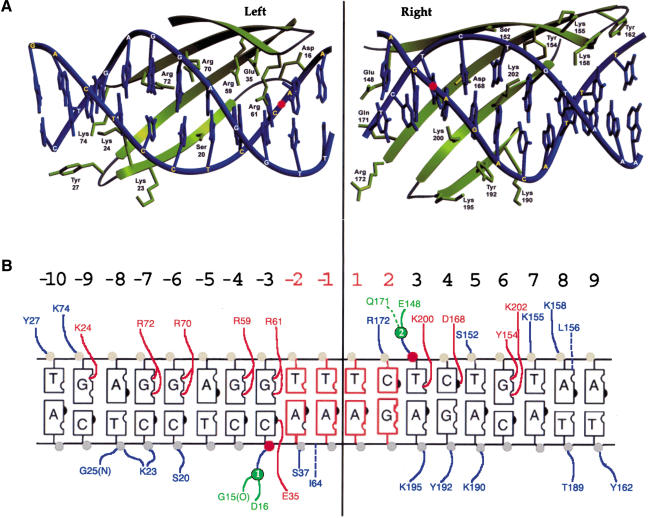
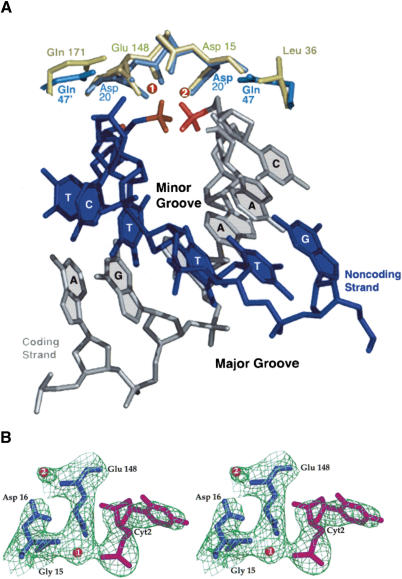
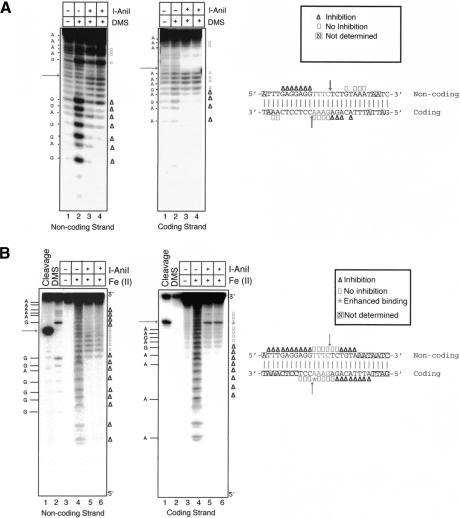
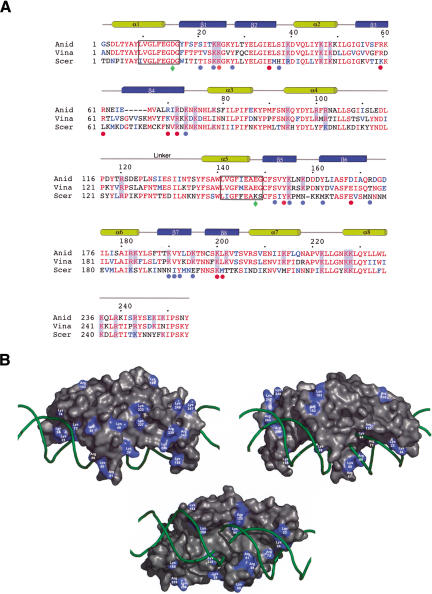
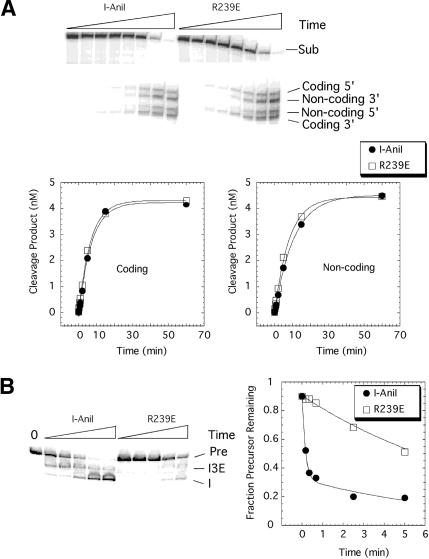
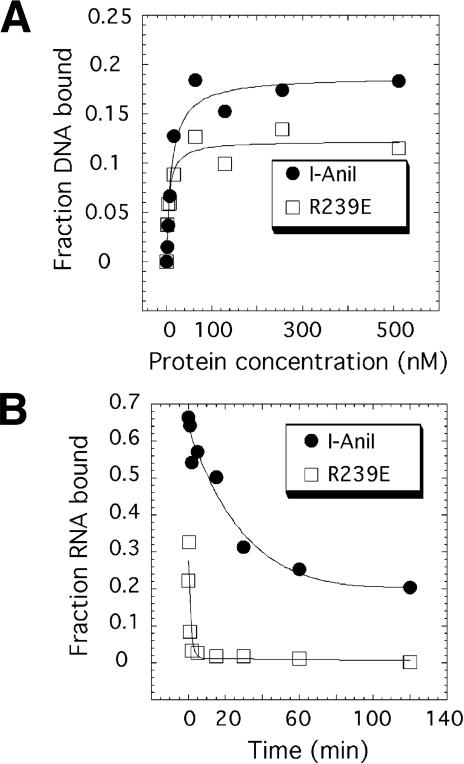
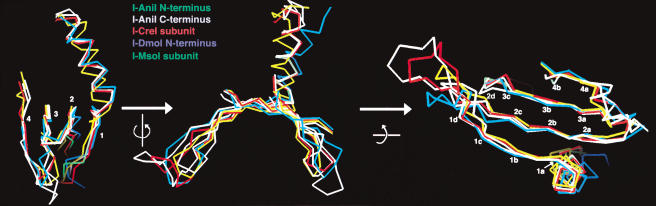
We determined the crystal structure of a bifunctional group I intron splicing factor and homing endonuclease, termed the I-AniI maturase, in complex with its DNA target at 2.6 A resolution. The structure demonstrates the remarkable structural conservation of the beta-sheet DNA-binding motif between highly divergent enzyme subfamilies. DNA recognition by I-AniI was further studied using nucleoside deletion and DMS modification interference analyses. Correlation of these results with the crystal structure provides information on the relative importance of individual nucleotide contacts for DNA recognition. Alignment and modeling of two homologous maturases reveals conserved basic surface residues, distant from the DNA-binding surface, that might be involved in RNA binding. A point mutation that introduces a single negative charge in this region uncouples the maturase and endonuclease functions of the protein, inhibiting RNA binding and splicing while maintaining DNA binding and cleavage.
Figures
Comment in
-
Two for the price of one: a bifunctional intron-encoded DNA endonuclease-RNA maturase.Genes Dev. 2003 Dec 1;17(23):2860-3. doi: 10.1101/gad.1162503. Genes Dev. 2003. PMID: 14665667 Review. No abstract available.
References
-
- Argast G.M., Stephens, K.M., Emond, M.J., and Monnat, R.J. 1998. I-PpoI and I-CreI homing site sequence degeneracy determined by random mutagenesis and sequential in vitro enrichment. J. Mol. Biol. 280: 345-353. - PubMed
-
- Belfort M. and Perlman, P.S. 1995. Mechanisms of intron mobility. J. Biol. Chem. 270: 30237-30240. - PubMed
-
- Brunger A. 1993. Assessment of phase accuracy by cross validation: The free R value. Methods and Applications. Acta Cryst. D49: 24-36. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases