

Hierarchical assembly of the budding yeast kinetochore from multiple subcomplexes
- PMID: 14633972
- PMCID: PMC289150
- DOI: 10.1101/gad.1144403
Hierarchical assembly of the budding yeast kinetochore from multiple subcomplexes
Abstract
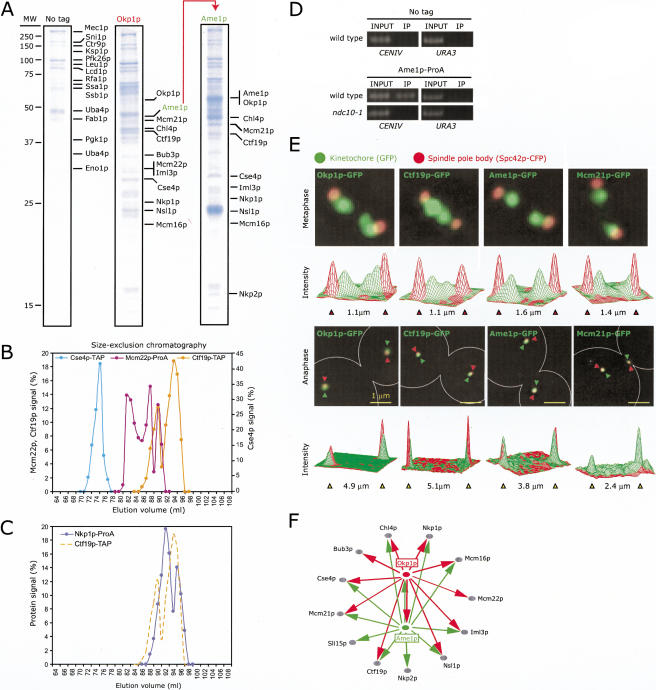
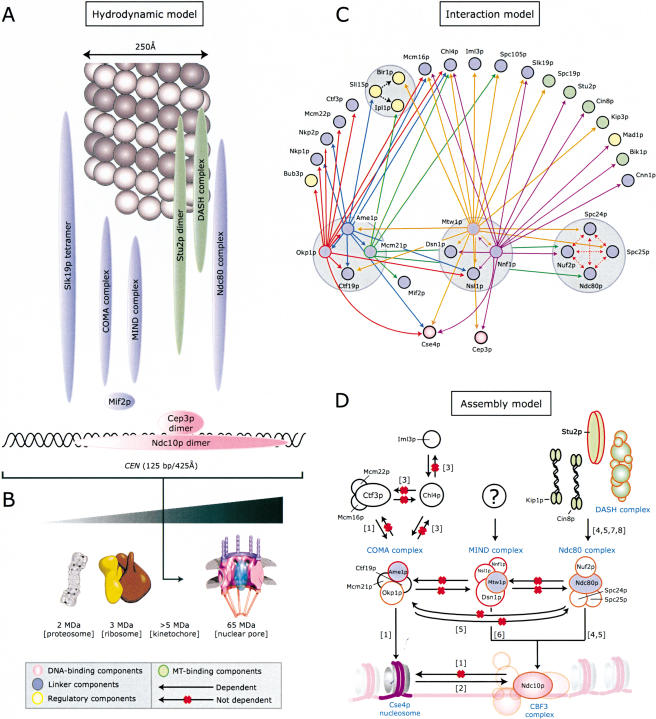
Kinetochores are multiprotein complexes that assemble on centromeric DNA and attach chromosomes to spindle microtubules. Over the past six years, the number of proteins known to localize to the Saccharomyces cerevisiae kinetochore has increased from around 10 to over 60. However, relatively little is known about the protein-protein interactions that mediate kinetochore assembly or about the overall structure of microtubule-attachment sites. Here we used biophysical techniques, affinity purification, mass spectrometry, and in vivo assays to examine the state of association of 31 centromere-binding proteins, including six proteins newly identified as kinetochore subunits. We found that yeast kinetochores resemble transcriptional enhancers in being composed of at least 17 discrete subcomplexes that assemble on DNA to form a very large structure with a mass in excess of 5 MD. Critical to kinetochore assembly are proteins that bridge subunits in direct contact with DNA and subunits bound to microtubules. We show that two newly identified kinetochore complexes, COMA (Ctf19p-Okp1p-Mcm21p-Ame1p) and MIND (Mtw1p including Nnf1p-Nsl1p-Dsn1p) function as bridges. COMA, MIND, and the previously described Ndc80 complex constitute three independent and essential platforms onto which outer kinetochore proteins assemble. In addition, we propose that the three complexes have different functions with respect to force generation and MT attachment.
Figures






References
-
- Bader G.D., Heilbut, A., Andrews, B., Tyers, M., Hughes, T., and Boone, C. 2003. Functional genomics and proteomics: Charting a multidimensional map of the yeast cell. Trends Cell. Biol. 13: 344-356. - PubMed
-
- Bell S.P. and Dutta, A. 2002. DNA replication in eukaryotic cells. Annu. Rev. Biochem. 71: 333-374. - PubMed
-
- Cheeseman I.M., Anderson, S., Jwa, M., Green, E.M., Kang, J., Yates 3rd, J.R., Chan, C.S., Drubin, D.G., and Barnes, G. 2002b. Phospho-regulation of kinetochore-microtubule attachments by the Aurora kinase Ipl1p. Cell 111: 163-172. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous