

Seasonal changes in the lower jaw skeleton in male Atlantic salmon (Salmo salar L.): remodelling and regression of the kype after spawning
- PMID: 14635799
- PMCID: PMC1571185
- DOI: 10.1046/j.1469-7580.2003.00239.x
Seasonal changes in the lower jaw skeleton in male Atlantic salmon (Salmo salar L.): remodelling and regression of the kype after spawning
Abstract
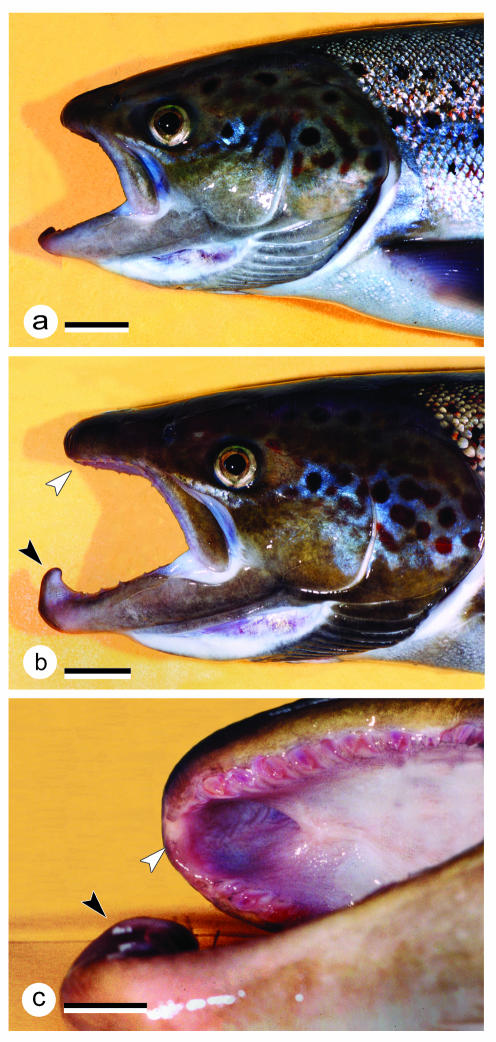
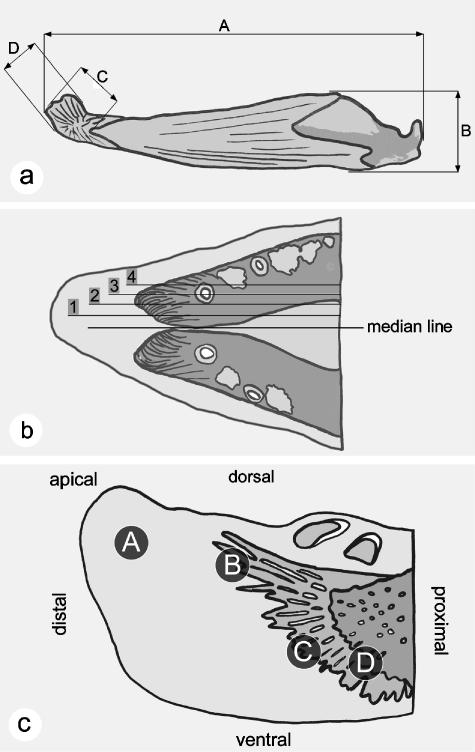
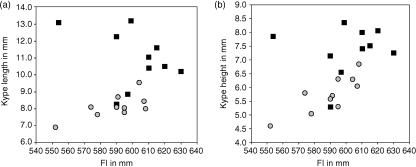
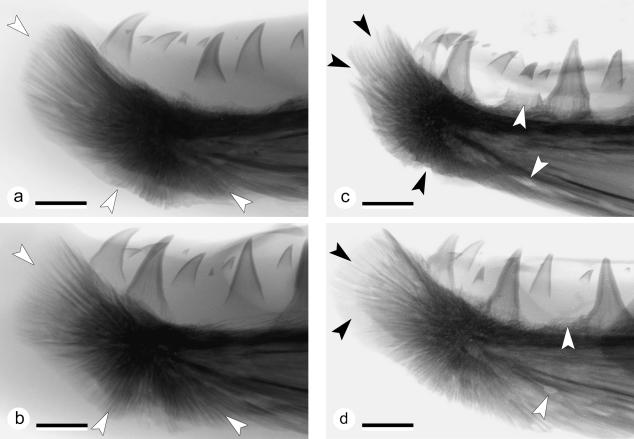
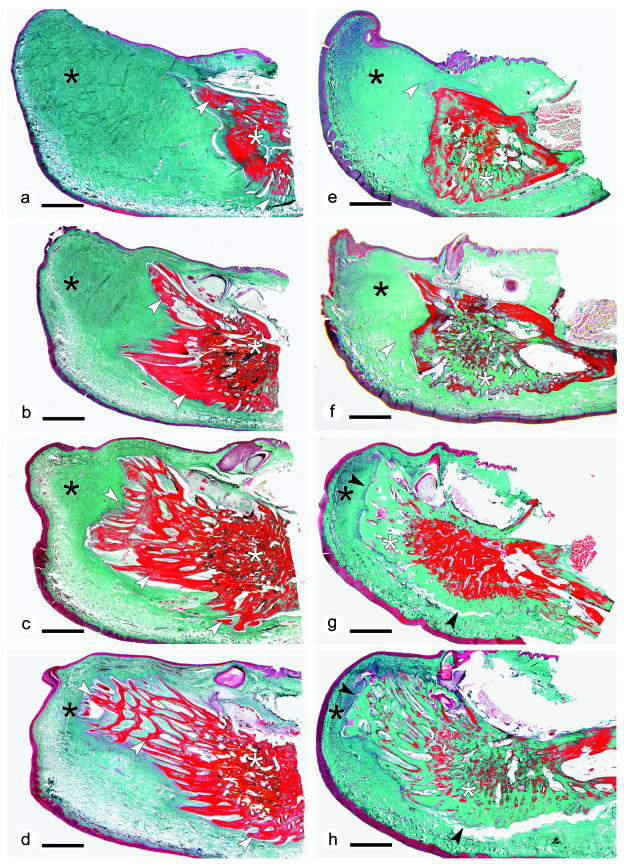
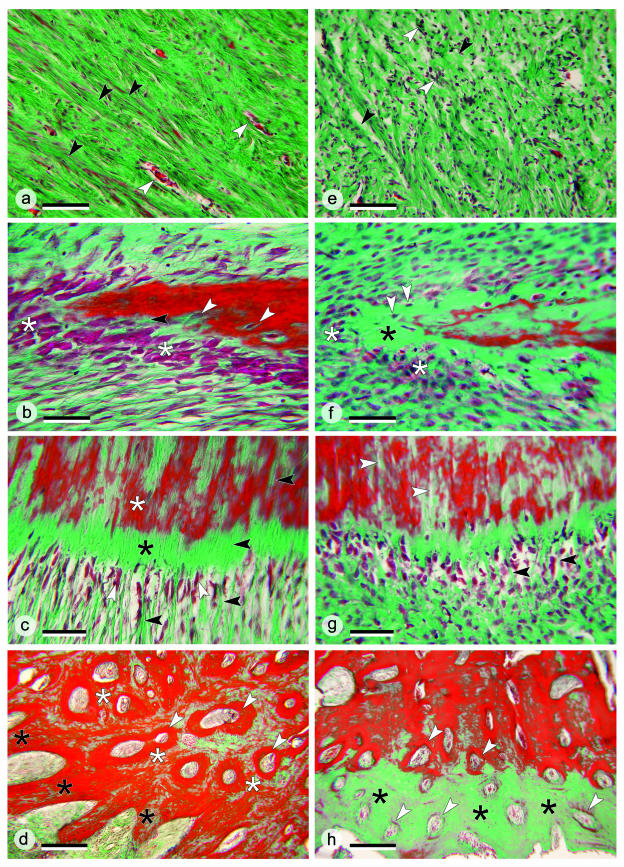
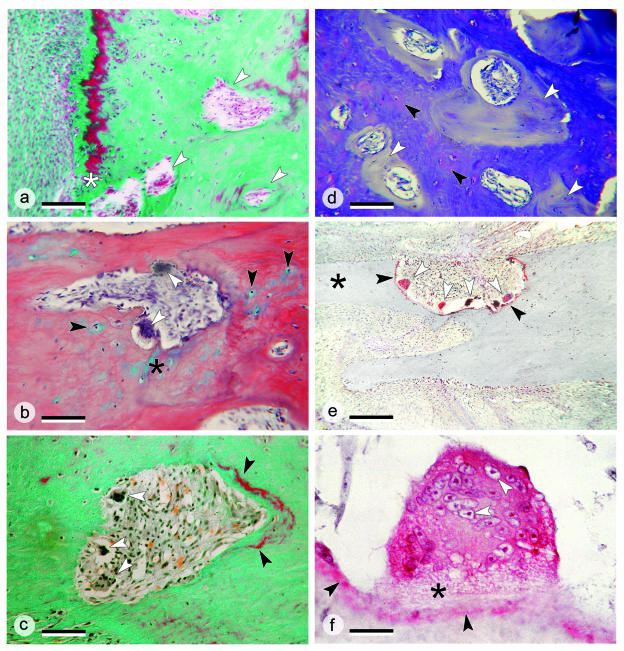
The return of Atlantic salmon (Salmon salar) to their home river for spawning coincides with drastic skeletal alterations in both sexes. Most prominent is the development of a kype (hook) at the tip of the lower jaw in males. Salmon that survive spawning have to cope with the kype throughout their life, unless it disappears after spawning, as was suggested in the early literature. To understand the fate of the kype skeleton, we compared morphological and histological features of kypes from pre-spawned mature anadromous males (grilse) with post-spawned males (kelts). The kype of male grilse is supported by fast-growing skeletal needles that differ from regular dentary bone. In kelts, growth of the kype skeleton has stopped and skeletal needles are resorbed apically by osteoclasts. Simultaneously, and despite the critical physiological condition of the animals, proximal parts of the kype skeleton are remodelled and converted into regular dentary bone. Apical resorption of the skeleton explains reports of a decrease of the kype in kelts. The conversion of basal kype skeleton into regular dentary bone contributes to the elongation of the dentary and probably also to the development of a larger kype in repetitive spawning males.
Figures
Similar articles
-
Differentiation and growth of kype skeletal tissues in anadromous male Atlantic salmon (Salmo salar).Int J Dev Biol. 2002 Aug;46(5):719-30. Int J Dev Biol. 2002. PMID: 12216984
-
Establishment, maintenance and modifications of the lower jaw dentition of wild Atlantic salmon (Salmo salar L.) throughout its life cycle.J Anat. 2007 Oct;211(4):471-84. doi: 10.1111/j.1469-7580.2007.00788.x. Epub 2007 Aug 31. J Anat. 2007. PMID: 17764526 Free PMC article.
-
Are breeding teeth in Atlantic salmon a component of the drastic alterations of the oral facial skeleton?Arch Oral Biol. 2005 Feb;50(2):213-7. doi: 10.1016/j.archoralbio.2004.10.018. Epub 2004 Dec 15. Arch Oral Biol. 2005. PMID: 15721152
-
A review of the likely effects of climate change on anadromous Atlantic salmon Salmo salar and brown trout Salmo trutta, with particular reference to water temperature and flow.J Fish Biol. 2009 Dec;75(10):2381-447. doi: 10.1111/j.1095-8649.2009.02380.x. J Fish Biol. 2009. PMID: 20738500 Review.
-
Temperature requirements of Atlantic salmon Salmo salar, brown trout Salmo trutta and Arctic charr Salvelinus alpinus: predicting the effects of climate change.J Fish Biol. 2010 Nov;77(8):1793-817. doi: 10.1111/j.1095-8649.2010.02762.x. Epub 2010 Oct 6. J Fish Biol. 2010. PMID: 21078091 Review.
Cited by
-
Matrix first, minerals later: fine-tuned dietary phosphate increases bone formation in zebrafish.JBMR Plus. 2024 Jun 18;8(8):ziae081. doi: 10.1093/jbmrpl/ziae081. eCollection 2024 Aug. JBMR Plus. 2024. PMID: 39045128 Free PMC article.
-
Evolutionary drivers of kype size in Atlantic salmon (Salmo salar): domestication, age and genetics.R Soc Open Sci. 2019 Apr 17;6(4):190021. doi: 10.1098/rsos.190021. eCollection 2019 Apr. R Soc Open Sci. 2019. PMID: 31183145 Free PMC article.
-
Embryonic origin of the gnathostome vertebral skeleton.Proc Biol Sci. 2017 Nov 29;284(1867):20172121. doi: 10.1098/rspb.2017.2121. Proc Biol Sci. 2017. PMID: 29167367 Free PMC article.
-
Evolutionary conservation and divergence of the transcriptional regulation of bivalve shell secretion across life-history stages.R Soc Open Sci. 2022 Dec 21;9(12):221022. doi: 10.1098/rsos.221022. eCollection 2022 Dec. R Soc Open Sci. 2022. PMID: 36569229 Free PMC article.
-
The pseudobranch of jawed vertebrates is a mandibular arch-derived gill.Development. 2022 Jul 1;149(13):dev200184. doi: 10.1242/dev.200184. Epub 2022 Jul 11. Development. 2022. PMID: 35762641 Free PMC article.
References
-
- Arndt SKA, Benfey TJ, Cunjak RA. Effect of temporary reductions in feeding on protein synthesis and energy storage of juvenile Atlantic salmon. J. Fish. Biol. 1996;49:257–276.
-
- Ashhurst DE. Macromolecular synthesis and mechanical stability during fracture repair. In: Hall BK, editor. Bone, Vol. 5: Fracture Repair and Regeneration. Boca Raton: CRC Press; 1992. pp. 61–121.
-
- Baker DM, Larsen DA, Swanson P, Dickhoff WW. Long-term peripheral treatment of immature coho salmon (Oncorhynchus kisutch) with human leptin has no clear physiologic effect. Gen. Comp. Endocrinol. 2000;118:134–138. - PubMed
-
- Benjamin M, Rufai A, Ralphs JR. The mechanism of formation of bony spurs (enthesophytes) in the Achilles tendon. Arthritis Rheum. 2000;43:576–583. - PubMed
-
- Beresford WA. Chondroid Bone, Secondary Cartilage and Metaplasia. Baltimore: Urban & Schwarzenberg; 1981.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
