

The Drosophila nucleoporin DNup88 localizes DNup214 and CRM1 on the nuclear envelope and attenuates NES-mediated nuclear export
- PMID: 14638854
- PMCID: PMC2173670
- DOI: 10.1083/jcb.200304046
The Drosophila nucleoporin DNup88 localizes DNup214 and CRM1 on the nuclear envelope and attenuates NES-mediated nuclear export
Abstract
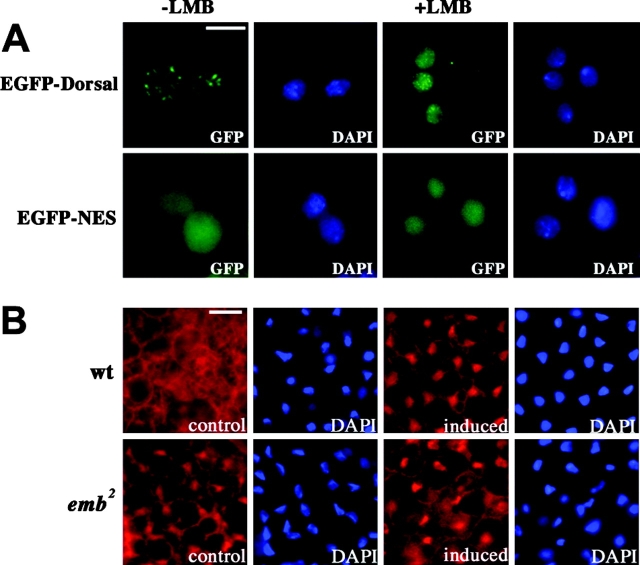
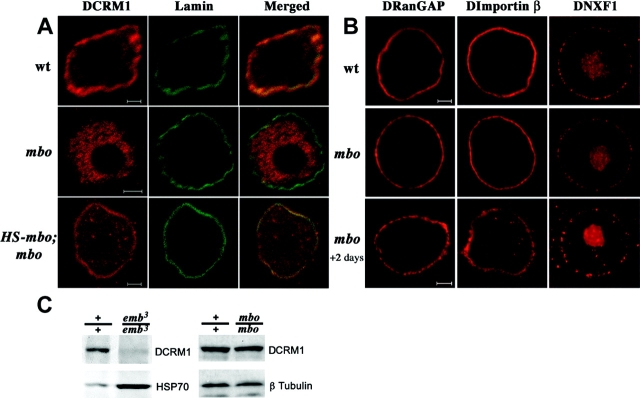
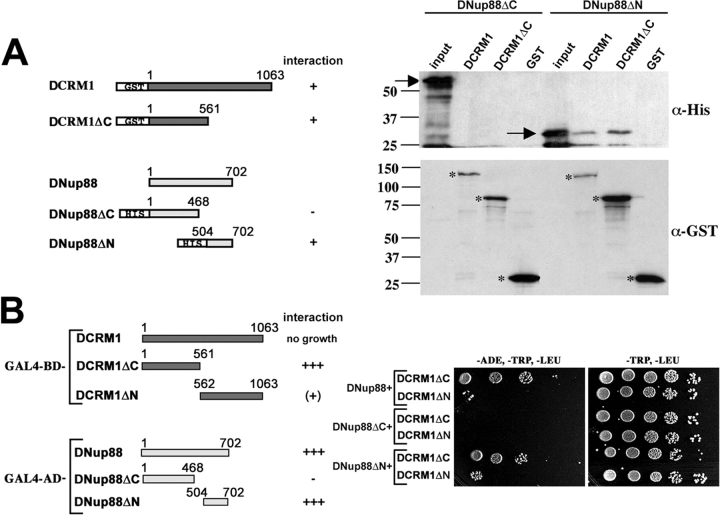
Many cellular responses rely on the control of nucleocytoplasmic transport of transcriptional regulators. The Drosophila nucleoporin Nup88 is selectively required for nuclear accumulation of Rel proteins and full activation of the innate immune response. Here, we investigate the mechanisms underlying its role in nucleocytoplasmic transport. Nuclear import of an nuclear localization signal-enhanced green fluorescent protein (NLS-EGFP) reporter is not affected in DNup88 (members only; mbo) mutants, whereas the level of CRM1-dependent EGFP-nuclear export signal (EGFP-NES) export is increased. We show that the nuclear accumulation of the Drosophila Rel protein Dorsal requires CRM1. DNup88 binds to DNup214 and DCRM1 in vitro, and both proteins become mislocalized from the nuclear rim into the nucleus of mbo mutants. Overexpression of DNup88 is sufficient to relocalize DNup214 and CRM1 on the nuclear envelope and revert the mutant phenotypes. We propose that a major function of DNup88 is to anchor DNup214 and CRM1 on the nuclear envelope and thereby attenuate NES-mediated nuclear export.
Figures


Similar articles
-
The nucleoporin Nup214 sequesters CRM1 at the nuclear rim and modulates NFkappaB activation in Drosophila.J Cell Sci. 2006 Nov 1;119(Pt 21):4409-19. doi: 10.1242/jcs.03201. Epub 2006 Oct 10. J Cell Sci. 2006. PMID: 17032737
-
The nucleoporin-like protein NLP1 (hCG1) promotes CRM1-dependent nuclear protein export.J Cell Sci. 2012 Jan 1;125(Pt 1):144-54. doi: 10.1242/jcs.090316. Epub 2012 Jan 16. J Cell Sci. 2012. PMID: 22250199
-
members only encodes a Drosophila nucleoporin required for rel protein import and immune response activation.Genes Dev. 2000 Aug 1;14(15):1945-57. Genes Dev. 2000. PMID: 10921908 Free PMC article.
-
Crossing the nuclear envelope: hierarchical regulation of nucleocytoplasmic transport.Science. 2007 Nov 30;318(5855):1412-6. doi: 10.1126/science.1142204. Science. 2007. PMID: 18048681 Review.
-
Nucleocytoplasmic transport of proteins.Biochemistry (Mosc). 2007 Dec;72(13):1439-57. doi: 10.1134/s0006297907130032. Biochemistry (Mosc). 2007. PMID: 18282135 Review.
Cited by
-
Mechanisms of Nuclear Export in Cancer and Resistance to Chemotherapy.Cancers (Basel). 2016 Mar 14;8(3):35. doi: 10.3390/cancers8030035. Cancers (Basel). 2016. PMID: 26985906 Free PMC article. Review.
-
Nucleo-cytoplasmic transport of proteins and RNA in plants.Plant Cell Rep. 2011 Feb;30(2):153-76. doi: 10.1007/s00299-010-0928-3. Epub 2010 Oct 20. Plant Cell Rep. 2011. PMID: 20960203 Free PMC article. Review.
-
Inhibition of CRM1-mediated nuclear export of transcription factors by leukemogenic NUP98 fusion proteins.J Biol Chem. 2010 May 21;285(21):16248-57. doi: 10.1074/jbc.M109.048785. Epub 2010 Mar 16. J Biol Chem. 2010. PMID: 20233715 Free PMC article.
-
Effects of gambogic acid on the regulation of nucleoporin Nup88 in U937 cells.J Huazhong Univ Sci Technolog Med Sci. 2007 Aug;27(4):388-92. doi: 10.1007/s11596-007-0410-9. J Huazhong Univ Sci Technolog Med Sci. 2007. PMID: 17828493
-
Central role of the oxygen-dependent degradation domain of Drosophila HIFalpha/Sima in oxygen-dependent nuclear export.Mol Biol Cell. 2009 Sep;20(17):3878-87. doi: 10.1091/mbc.e09-01-0038. Epub 2009 Jul 8. Mol Biol Cell. 2009. PMID: 19587118 Free PMC article.
References
-
- Ausubel, F.M., R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman, J.A. Smith, and K. Struhl, editors. 1993. Calcium phosphate transfection. In Current Protocols in Molecular Biology. John Wiley & Sons, Inc., New York. 9.1.1–9.1.4.
-
- Bergmann, A., D. Stein, R. Geisler, S. Hagenmaier, B. Schmid, N. Fernandez, B. Schnell, and C. Nusslein-Volhard. 1996. A gradient of cytoplasmic Cactus degradation establishes the nuclear localization gradient of the dorsal morphogen in Drosophila. Mech. Dev. 60:109–123. - PubMed
-
- Carlotti, F., S.K. Dower, and E.E. Qwarnstrom. 2000. Dynamic shuttling of nuclear factor kappa B between the nucleus and cytoplasm as a consequence of inhibitor dissociation. J. Biol. Chem. 275:41028–41034. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
