

Interactions between electron and proton currents in excised patches from human eosinophils
- PMID: 14638931
- PMCID: PMC2229590
- DOI: 10.1085/jgp.200308891
Interactions between electron and proton currents in excised patches from human eosinophils
Abstract
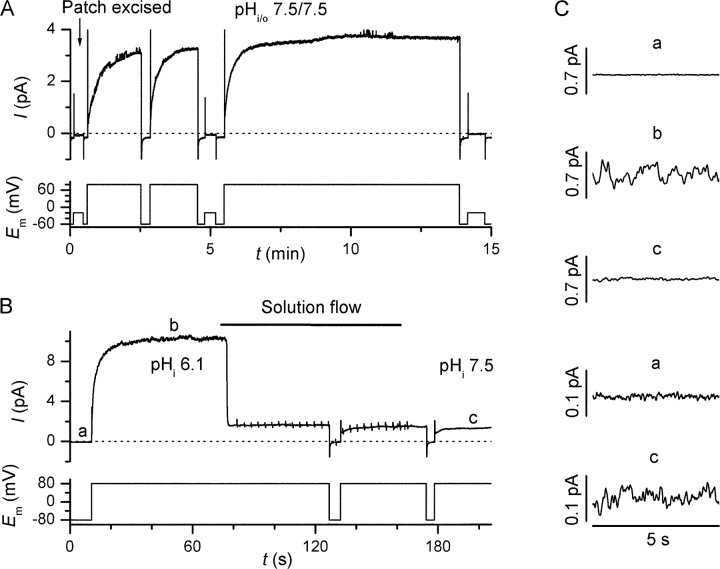
The NADPH-oxidase is a plasma membrane enzyme complex that enables phagocytes to generate superoxide in order to kill invading pathogens, a critical step in the host defense against infections. The oxidase transfers electrons from cytosolic NADPH to extracellular oxygen, a process that requires concomitant H+ extrusion through depolarization-activated H+ channels. Whether H+ fluxes are mediated by the oxidase itself is controversial, but there is a general agreement that the oxidase and H+ channel are intimately connected. Oxidase activation evokes profound changes in whole-cell H+ current (IH), causing an approximately -40-mV shift in the activation threshold that leads to the appearance of inward IH. To further explore the relationship between the oxidase and proton channel, we performed voltage-clamp experiments on inside-out patches from both resting and phorbol-12-myristate-13-acetate (PMA)-activated human eosinophils. Proton currents from resting cells displayed slow voltage-dependent activation, long-term stability, and were blocked by micromolar internal [Zn2+]. IH from PMA-treated cells activated faster and at lower voltages, enabling sustained H+ influx, but ran down within minutes, regaining the current properties of nonactivated cells. Bath application of NADPH to patches excised from PMA-treated cells evoked electron currents (Ie), which also ran down within minutes and were blocked by diphenylene iodonium (DPI). Run-down of both IH and Ie was delayed, and sometimes prevented, by cytosolic ATP and GTP-gamma-S. A good correlation was observed between the amplitude of Ie and both inward and outward IH when a stable driving force for e- was imposed. Combined application of NADPH and DPI reduced the inward IH amplitude, even in the absence of concomitant oxidase activity. The strict correlation between Ie and IH amplitudes and the sensitivity of IH to oxidase-specific agents suggest that the proton channel is either part of the oxidase complex or linked by a membrane-limited mediator.
Figures







Similar articles
-
Interactions between NADPH oxidase-related proton and electron currents in human eosinophils.J Physiol. 2001 Sep 15;535(Pt 3):767-81. doi: 10.1111/j.1469-7793.2001.00767.x. J Physiol. 2001. PMID: 11559774 Free PMC article.
-
Regulation of eosinophil membrane depolarization during NADPH oxidase activation.J Cell Sci. 2003 Aug 1;116(Pt 15):3221-6. doi: 10.1242/jcs.00627. J Cell Sci. 2003. PMID: 12829741
-
Electron and proton transport by NADPH oxidases.Philos Trans R Soc Lond B Biol Sci. 2005 Dec 29;360(1464):2315-25. doi: 10.1098/rstb.2005.1769. Philos Trans R Soc Lond B Biol Sci. 2005. PMID: 16321802 Free PMC article. Review.
-
Simultaneous activation of NADPH oxidase-related proton and electron currents in human neutrophils.Proc Natl Acad Sci U S A. 2000 Jun 6;97(12):6885-9. doi: 10.1073/pnas.100047297. Proc Natl Acad Sci U S A. 2000. PMID: 10823889 Free PMC article.
-
NADPH oxidase subunit gp91phox: a proton pathway.Protoplasma. 2001;217(1-3):37-42. doi: 10.1007/BF01289411. Protoplasma. 2001. PMID: 11732336 Review.
Cited by
-
Consequences of the electrogenic function of the phagocytic NADPH oxidase.Philos Trans R Soc Lond B Biol Sci. 2005 Dec 29;360(1464):2293-300. doi: 10.1098/rstb.2005.1768. Philos Trans R Soc Lond B Biol Sci. 2005. PMID: 16321799 Free PMC article. Review.
-
Molecular mechanisms of coupling to voltage sensors in voltage-evoked cellular signals.Proc Jpn Acad Ser B Phys Biol Sci. 2019;95(3):111-135. doi: 10.2183/pjab.95.010. Proc Jpn Acad Ser B Phys Biol Sci. 2019. PMID: 30853698 Free PMC article. Review.
-
Voltage-gated proton channels: what's next?J Physiol. 2008 Nov 15;586(22):5305-24. doi: 10.1113/jphysiol.2008.161703. Epub 2008 Sep 18. J Physiol. 2008. PMID: 18801839 Free PMC article. Review.
-
The pH dependence of NADPH oxidase in human eosinophils.J Physiol. 2005 Dec 1;569(Pt 2):419-31. doi: 10.1113/jphysiol.2005.094748. Epub 2005 Sep 29. J Physiol. 2005. PMID: 16195320 Free PMC article.
-
Voltage-Gated Proton Channels as Novel Drug Targets: From NADPH Oxidase Regulation to Sperm Biology.Antioxid Redox Signal. 2015 Aug 10;23(5):490-513. doi: 10.1089/ars.2013.5806. Epub 2014 Mar 17. Antioxid Redox Signal. 2015. PMID: 24483328 Free PMC article. Review.
References
-
- Babior, B.M., J.D. Lambeth, and W. Nauseef. 2002. The neutrophil NADPH oxidase. Arch. Biochem. Biophys. 397:342–344. - PubMed
-
- Banfi, B., A. Maturana, S. Jaconi, S. Arnaudeau, T. Laforge, B. Sinha, E. Ligeti, N. Demaurex, and K.H. Krause. 2000. A mammalian H+ channel generated through alternative splicing of the NADPH oxidase homolog NOH-1. Science. 287:138–142. - PubMed
-
- Banfi, B., G. Molnar, A. Maturana, K. Steger, B. Hegedus, N. Demaurex, and K.H. Krause. 2001. A Ca2+-activated NADPH oxidase in testis, spleen, and lymph nodes. J. Biol. Chem. 276:37594–37601. - PubMed
-
- Biberstine-Kinkade, K.J., F.R. DeLeo, R.I. Epstein, B.A. LeRoy, W.M. Nauseef, and M.C. Dinauer. 2001. Heme-ligating histidines in flavocytochrome b(558): identification of specific histidines in gp91(phox). J. Biol. Chem. 276:31105–31112. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
