

Developmentally specific role of the CCAAT box in regulation of human gamma-globin gene expression
- PMID: 14645237
- PMCID: PMC2808414
- DOI: 10.1074/jbc.M306241200
Developmentally specific role of the CCAAT box in regulation of human gamma-globin gene expression
Abstract
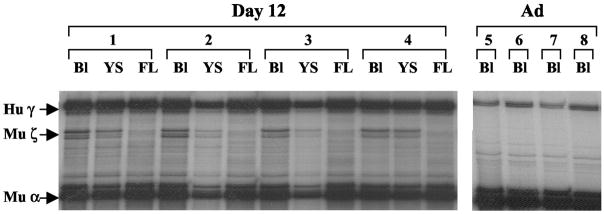
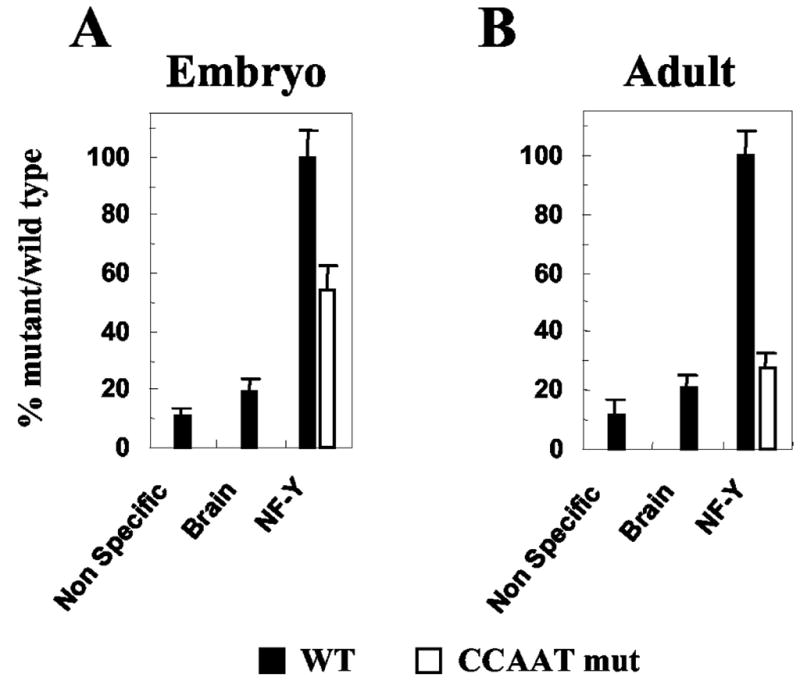
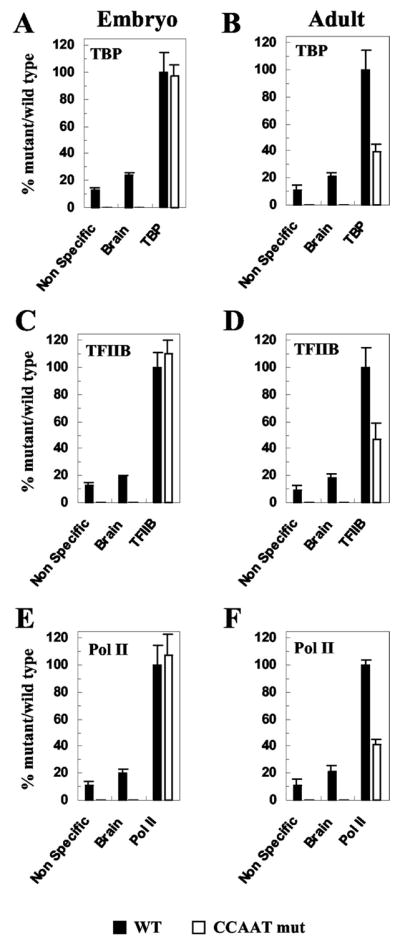
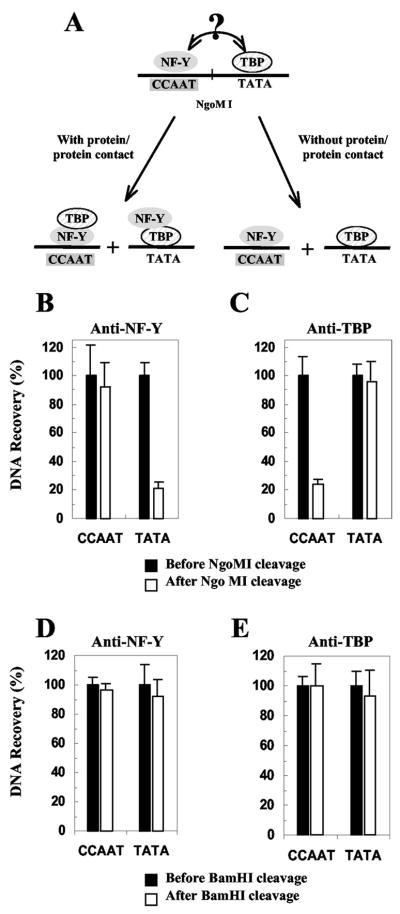
The CCAAT box is a widespread motif in eukaryotic promoters. In this study we demonstrate that the effects of the CCAAT box on gamma-globin gene activation are developmentally distinct. Although this promoter element is essential for high level gamma gene expression in adult erythropoiesis, it plays little role in embryonic erythroid cells. The CCAAT mutation in the human gamma-globin gene promoter impairs recruitment of TATA-binding protein (TBP), TFIIB, and RNA polymerase II in adult splenic erythroblasts but not in embryonic erythroid cells. We also show that the efficiency of gamma gene transcription is correlated with recruitment of TBP on the TATA box but that the level of TBP recruitment is not nuclear factor Y (NF-Y)-dependent. Our data also suggest that it is unlikely that transcriptional stimulation by the CCAAT box is exerted through direct protein-protein interaction between NF-Y and TBP.
Figures
Similar articles
-
Developmental specificity of recruitment of TBP to the TATA box of the human gamma-globin gene.Proc Natl Acad Sci U S A. 2002 Apr 16;99(8):5509-14. doi: 10.1073/pnas.072084499. Proc Natl Acad Sci U S A. 2002. PMID: 11960008 Free PMC article.
-
Transcriptional potential of the gamma-globin gene is dependent on the CACCC box in a developmental stage-specific manner.Nucleic Acids Res. 2006;34(14):3909-16. doi: 10.1093/nar/gkl474. Epub 2006 Aug 12. Nucleic Acids Res. 2006. PMID: 16914453 Free PMC article.
-
The regulation of human globin promoters by CCAAT box elements and the recruitment of NF-Y.Biochim Biophys Acta Gene Regul Mech. 2017 May;1860(5):525-536. doi: 10.1016/j.bbagrm.2016.10.002. Epub 2016 Oct 5. Biochim Biophys Acta Gene Regul Mech. 2017. PMID: 27718361 Review.
-
NF-Y recruits both transcription activator and repressor to modulate tissue- and developmental stage-specific expression of human γ-globin gene.PLoS One. 2012;7(10):e47175. doi: 10.1371/journal.pone.0047175. Epub 2012 Oct 10. PLoS One. 2012. PMID: 23071749 Free PMC article.
-
Role of the TATA-box binding protein (TBP) and associated family members in transcription regulation.Gene. 2022 Jul 30;833:146581. doi: 10.1016/j.gene.2022.146581. Epub 2022 May 18. Gene. 2022. PMID: 35597524 Review.
Cited by
-
Transcription factor competition at the γ-globin promoters controls hemoglobin switching.Nat Genet. 2021 Apr;53(4):511-520. doi: 10.1038/s41588-021-00798-y. Epub 2021 Mar 1. Nat Genet. 2021. PMID: 33649594 Free PMC article.
-
Inhibition of CBF/NF-Y mediated transcription activation arrests cells at G2/M phase and suppresses expression of genes activated at G2/M phase of the cell cycle.Nucleic Acids Res. 2006;34(21):6272-85. doi: 10.1093/nar/gkl801. Epub 2006 Nov 10. Nucleic Acids Res. 2006. PMID: 17098936 Free PMC article.
-
Phylogenetic comparisons suggest that distance from the locus control region guides developmental expression of primate beta-type globin genes.Proc Natl Acad Sci U S A. 2006 Feb 28;103(9):3186-91. doi: 10.1073/pnas.0511347103. Epub 2006 Feb 17. Proc Natl Acad Sci U S A. 2006. PMID: 16488971 Free PMC article.
-
TAT-mediated transduction of NF-Ya peptide induces the ex vivo proliferation and engraftment potential of human hematopoietic progenitor cells.Blood. 2010 Oct 14;116(15):2676-83. doi: 10.1182/blood-2010-03-273441. Epub 2010 Jul 8. Blood. 2010. PMID: 20616221 Free PMC article.
-
Gene expression profiling identifies HOXB4 as a direct downstream target of GATA-2 in human CD34+ hematopoietic cells.PLoS One. 2012;7(9):e40959. doi: 10.1371/journal.pone.0040959. Epub 2012 Sep 24. PLoS One. 2012. PMID: 23028422 Free PMC article.
References
-
- Bucher P. J Mol Biol. 1990;212:563–578. - PubMed
-
- Mantovani R. Gene (Amst) 1999;239:15–27. - PubMed
-
- Maire P, Wuarin J, Schibler U. Science. 1989;244:343–346. - PubMed
-
- Jackson SM, Ericsson J, Osborne TF, Edwards PA. J Biol Chem. 1995;270:21445–21448. - PubMed
-
- Hsiao KM, McMahon SL, Farnham PJ. Genes Dev. 1994;8:1526–1537. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
