

Human beta-globin locus control region HS5 contains CTCF- and developmental stage-dependent enhancer-blocking activity in erythroid cells
- PMID: 14645507
- PMCID: PMC309639
- DOI: 10.1128/MCB.23.24.8946-8952.2003
Human beta-globin locus control region HS5 contains CTCF- and developmental stage-dependent enhancer-blocking activity in erythroid cells
Abstract
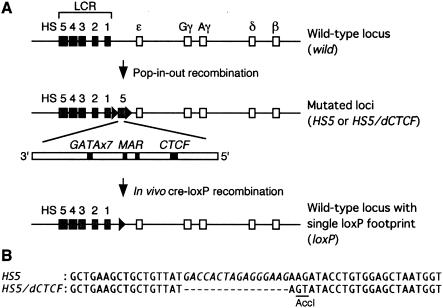
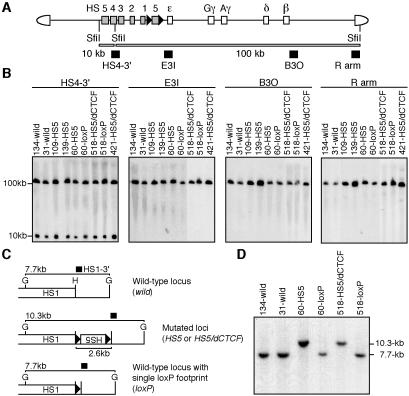
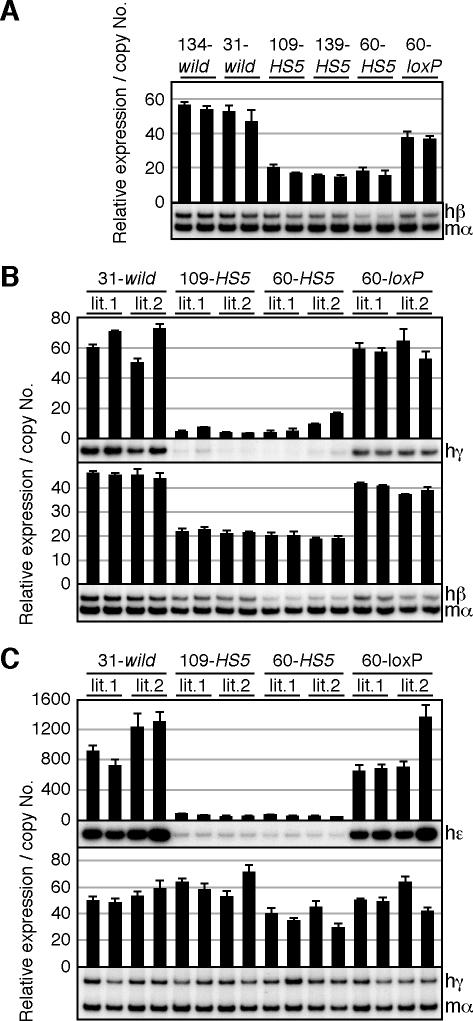
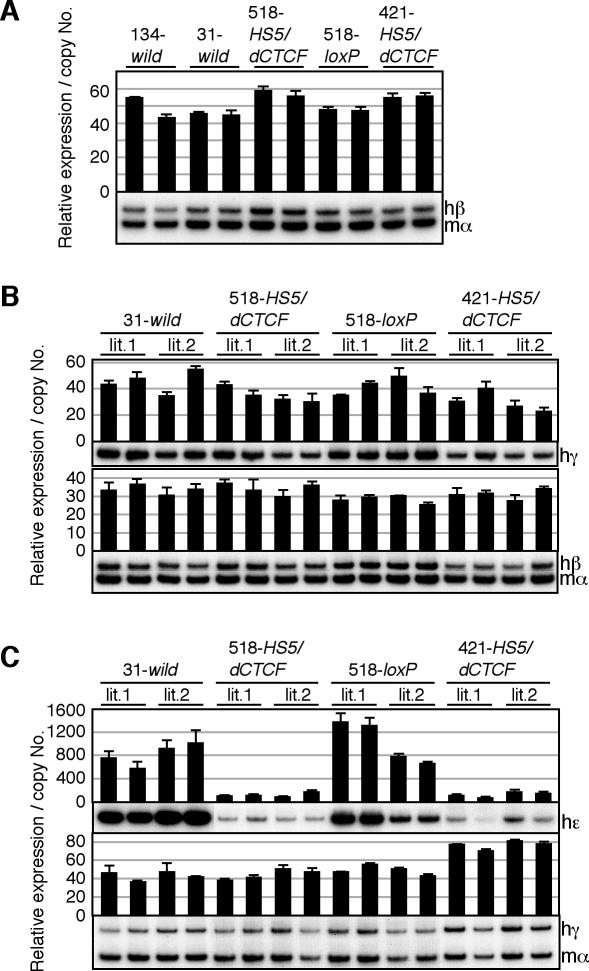
The human beta-globin locus contains five developmentally regulated beta-type globin genes. All five genes depend on the locus control region (LCR), located at the 5' end of the locus, for abundant globin gene transcription. The LCR is composed of five DNase I-hypersensitive sites (HSs), at least a subset of which appear to cooperate to form a holocomplex in activating genes within the locus. We previously tested the requirement for proper LCR polarity by inverting it in human beta-globin yeast artificial chromosome transgenic mice and observed reduced expression of all the beta-type globin genes regardless of developmental stage. This phenotype clearly demonstrated an orientation-dependent activity of the LCR, although the mechanistic basis for the observed activity was obscure. Here, we describe genetic evidence demonstrating that human HS5 includes enhancer-blocking (insulator) activity that is both CTCF and developmental stage dependent. Curiously, we also observed an attenuating activity in HS5 that was specific to the epsilon-globin gene at the primitive stage and was independent of the HS5 CTCF binding site. These observations demonstrate that the phenotype observed in the LCR-inverted locus was in part attributable to placing the HS5 insulator between the LCR HS enhancers (HS1 to HS4) and the promoter of the beta-globin gene.
Figures
Similar articles
-
DNase I hypersensitivity and epsilon-globin transcriptional enhancement are separable in locus control region (LCR) HS1 mutant human beta-globin YAC transgenic mice.J Biol Chem. 2010 May 7;285(19):14495-503. doi: 10.1074/jbc.M110.116525. Epub 2010 Mar 15. J Biol Chem. 2010. PMID: 20231293 Free PMC article.
-
CTCF-dependent enhancer-blocking by alternative chromatin loop formation.Proc Natl Acad Sci U S A. 2008 Dec 23;105(51):20398-403. doi: 10.1073/pnas.0808506106. Epub 2008 Dec 12. Proc Natl Acad Sci U S A. 2008. PMID: 19074263 Free PMC article.
-
Conserved CTCF insulator elements flank the mouse and human beta-globin loci.Mol Cell Biol. 2002 Jun;22(11):3820-31. doi: 10.1128/MCB.22.11.3820-3831.2002. Mol Cell Biol. 2002. PMID: 11997516 Free PMC article.
-
beta-YAC transgenic mice for studying LCR function.Ann N Y Acad Sci. 1998 Jun 30;850:28-37. doi: 10.1111/j.1749-6632.1998.tb10459.x. Ann N Y Acad Sci. 1998. PMID: 9668524 Review.
-
Chromatin looping as a target for altering erythroid gene expression.Ann N Y Acad Sci. 2016 Mar;1368(1):31-9. doi: 10.1111/nyas.13012. Epub 2016 Feb 25. Ann N Y Acad Sci. 2016. PMID: 26918894 Free PMC article. Review.
Cited by
-
Creating New β-Globin-Expressing Lentiviral Vectors by High-Resolution Mapping of Locus Control Region Enhancer Sequences.Mol Ther Methods Clin Dev. 2020 Apr 18;17:999-1013. doi: 10.1016/j.omtm.2020.04.006. eCollection 2020 Jun 12. Mol Ther Methods Clin Dev. 2020. PMID: 32426415 Free PMC article.
-
Kaposi's Sarcoma-Associated Herpesvirus Genome Replication, Partitioning, and Maintenance in Latency.Front Microbiol. 2012 Jan 24;3:7. doi: 10.3389/fmicb.2012.00007. eCollection 2012. Front Microbiol. 2012. PMID: 22291692 Free PMC article.
-
Evaluation of regulatory potential and conservation scores for detecting cis-regulatory modules in aligned mammalian genome sequences.Genome Res. 2005 Aug;15(8):1051-60. doi: 10.1101/gr.3642605. Epub 2005 Jul 15. Genome Res. 2005. PMID: 16024817 Free PMC article.
-
The chicken HS4 insulator element does not protect the H19 ICR from differential DNA methylation in yeast artificial chromosome transgenic mouse.PLoS One. 2013 Sep 4;8(9):e73925. doi: 10.1371/journal.pone.0073925. eCollection 2013. PLoS One. 2013. PMID: 24023912 Free PMC article.
-
An insulator with barrier-element activity promotes alpha-spectrin gene expression in erythroid cells.Blood. 2009 Feb 12;113(7):1547-54. doi: 10.1182/blood-2008-06-164954. Epub 2008 Nov 13. Blood. 2009. PMID: 19008453 Free PMC article.
References
-
- Bell, A. C., A. G. West, and G. Felsenfeld. 1999. The protein CTCF is required for the enhancer-blocking activity of vertebrate insulators. Cell 98:387-396. - PubMed
-
- Bender, M. A., A. Reik, J. Close, A. Telling, E. Epner, S. Fiering, R. Hardison, and M. Groudine. 1998. Description and targeted deletion of 5′ hypersensitive site 5 and 6 of the mouse beta-globin locus control region. Blood 92:4394-4403. - PubMed
-
- Bulger, M., M. A. Bender, J. H. van Doorninck, B. Wertman, C. M. Farrell, G. Felsenfeld, M. Groudine, and R. Hardison. 2000. Comparative structural and functional analysis of the olfactory receptor genes flanking the human and mouse beta-globin gene clusters. Proc. Natl. Acad. Sci. USA 97:14560-14565. - PMC - PubMed
-
- Bulger, M., J. H. van Doorninck, N. Saitoh, A. Telling, C. Farrell, M. A. Bender, G. Felsenfeld, R. Axel, M. Groudine, and J. H. von Doorninck. 1999. Conservation of sequence and structure flanking the mouse and human beta-globin loci: the beta-globin genes are embedded within an array of odorant receptor genes. Proc. Natl. Acad. Sci. USA 96:5129-5134. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous