

Chromosomal integration and homologous gene targeting by replication-incompetent vectors based on the autonomous parvovirus minute virus of mice
- PMID: 14645570
- PMCID: PMC296056
- DOI: 10.1128/jvi.77.24.13136-13145.2003
Chromosomal integration and homologous gene targeting by replication-incompetent vectors based on the autonomous parvovirus minute virus of mice
Abstract
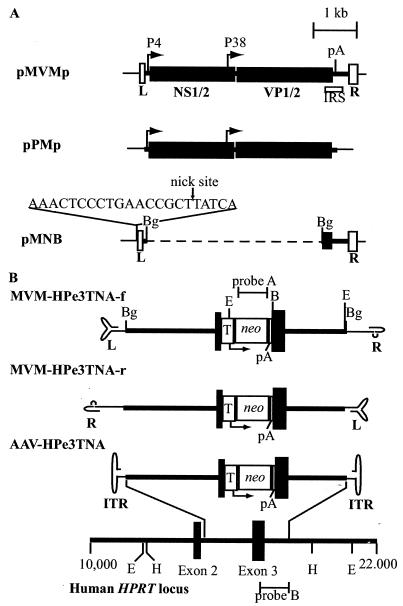
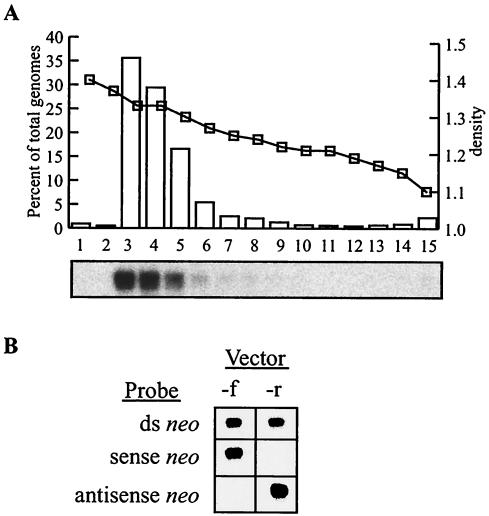
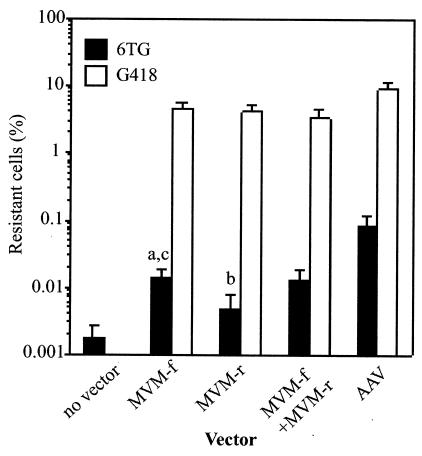
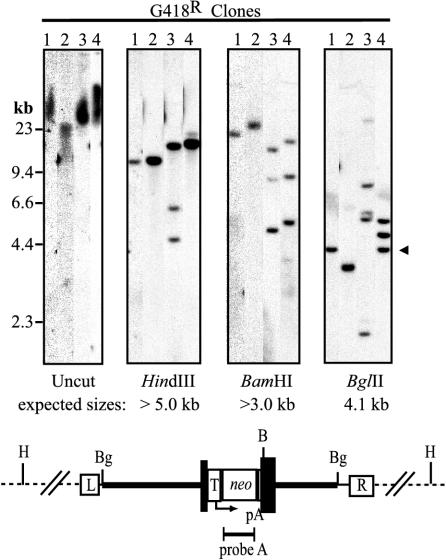
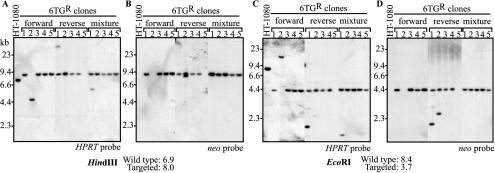
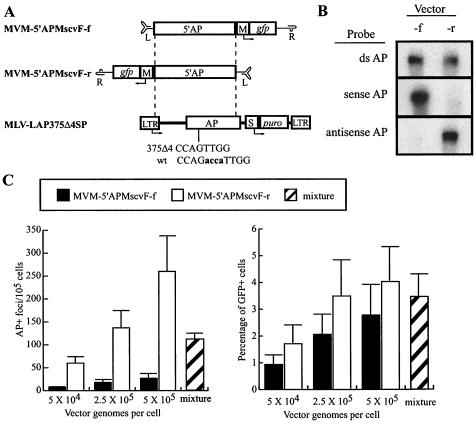
The molecular mechanisms responsible for random integration and gene targeting by recombinant adeno-associated virus (AAV) vectors are largely unknown, and whether vectors derived from autonomous parvoviruses transduce cells by similar pathways has not been investigated. In this report, we constructed vectors based on the autonomous parvovirus minute virus of mice (MVM) that were designed to introduce a neomycin resistance expression cassette (neo) into the X-linked human hypoxanthine phosphoribosyl transferase (HPRT) locus. High-titer, replication-incompetent MVM vector stocks were generated with a two-plasmid transfection system that preserved the wild-type characteristic of packaging only one DNA strand. Vectors with inserts in the forward or reverse orientations packaged noncoding or coding strands, respectively. In human HT-1080 cells, MVM vector random integration frequencies (neo(+) colonies) were comparable to those obtained with AAV vectors, and no difference was observed for noncoding and coding strands. HPRT gene-targeting frequencies (HPRT mutant colonies) were lower with MVM vectors, and the noncoding strand frequency was threefold greater than that of the coding strand. Random integration and gene-targeting events were confirmed by Southern blot analysis of G418- and 6-thioguanine (6TG)-resistant clones. In separate experiments, correction of an alkaline phosphatase (AP) gene by gene targeting was nine times more effective with a coding strand vector. The data suggest that single-stranded parvoviral vector genomes are substrates for gene targeting and possibly for random integration as well.
Figures


Similar articles
-
Chromosomal latency and expression at map unit 96 of a wild-type plus adeno-associated virus (AAV)/Neo vector and identification of p81, a new AAV transcriptional promoter.J Hum Virol. 1999 Nov-Dec;2(6):359-68. J Hum Virol. 1999. PMID: 10774553
-
Construction and production of oncotropic vectors, derived from MVM(p), that share reduced sequence homology with helper plasmids.Cancer Gene Ther. 2002 Sep;9(9):762-70. doi: 10.1038/sj.cgt.7700496. Cancer Gene Ther. 2002. PMID: 12189526
-
Influence of sequence and size of DNA on packaging efficiency of parvovirus MVM-based vectors.Hum Gene Ther. 1999 May 1;10(7):1229-38. doi: 10.1089/10430349950018210. Hum Gene Ther. 1999. PMID: 10340554
-
Autonomous parvovirus vectors: preventing the generation of wild-type or replication-competent virus.J Gene Med. 2004 Feb;6 Suppl 1:S203-11. doi: 10.1002/jgm.504. J Gene Med. 2004. PMID: 14978763 Review.
-
Parvoviral vectors for the gene therapy of cancer.Semin Oncol. 1996 Feb;23(1):159-71. Semin Oncol. 1996. PMID: 8607026 Review.
Cited by
-
A crisper look at genome editing: RNA-guided genome modification.Mol Ther. 2013 Apr;21(4):720-2. doi: 10.1038/mt.2013.46. Mol Ther. 2013. PMID: 23542565 Free PMC article. No abstract available.
-
AAV-mediated gene targeting is significantly enhanced by transient inhibition of nonhomologous end joining or the proteasome in vivo.Hum Gene Ther. 2012 Jun;23(6):658-65. doi: 10.1089/hum.2012.038. Epub 2012 Jun 25. Hum Gene Ther. 2012. PMID: 22486314 Free PMC article.
-
The effects of polymorphisms on human gene targeting.Nucleic Acids Res. 2014 Mar;42(5):3119-24. doi: 10.1093/nar/gkt1303. Epub 2013 Dec 25. Nucleic Acids Res. 2014. PMID: 24371280 Free PMC article.
-
Oligo/polynucleotide-based gene modification: strategies and therapeutic potential.Oligonucleotides. 2011 Mar-Apr;21(2):55-75. doi: 10.1089/oli.2010.0273. Epub 2011 Mar 21. Oligonucleotides. 2011. PMID: 21417933 Free PMC article. Review.
-
Inactivation of hepatitis B virus replication in cultured cells and in vivo with engineered transcription activator-like effector nucleases.Mol Ther. 2013 Oct;21(10):1889-97. doi: 10.1038/mt.2013.170. Epub 2013 Jul 25. Mol Ther. 2013. PMID: 23883864 Free PMC article.
References
-
- Aitken, M. L., R. B. Moss, D. A. Waltz, M. E. Dovey, M. R. Tonelli, S. C. McNamara, R. L. Gibson, B. W. Ramsey, B. J. Carter, and T. C. Reynolds. 2001. A phase I study of aerosolized administration of tgAAVCF to cystic fibrosis subjects with mild lung disease. Hum. Gene Ther. 12:1907-1916. - PubMed
-
- Avalosse, B. L., F. Dupont, P. Spegelaere, N. Mine, and A. Burny. 1996. Method for concentrating and purifying recombinant autonomous parvovirus vectors designed for tumour-cell-targeted gene therapy. J. Virol. Methods 62:179-183. - PubMed
-
- Berns, K. I. 1996. Parvoviridae: the viruses and their replication, p. 2173-2197. In B. N. Fields, D. M. Knipe, P. M. Howley et al. (ed.), Fields virology, 3rd ed. Lippincott-Raven Publishers, Philadelphia, Pa.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
