

Requirement of p53 targets in chemosensitization of colonic carcinoma to death ligand therapy
- PMID: 14645705
- PMCID: PMC299914
- DOI: 10.1073/pnas.2435285100
Requirement of p53 targets in chemosensitization of colonic carcinoma to death ligand therapy
Abstract
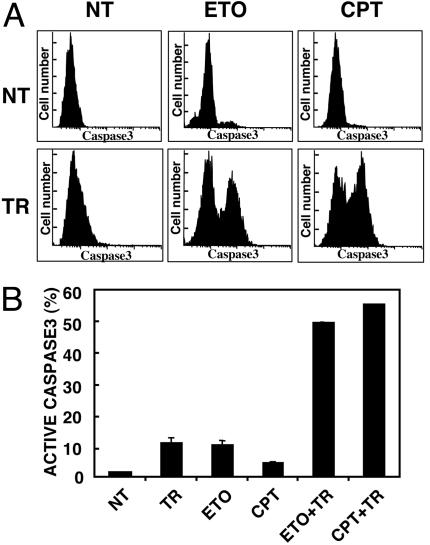
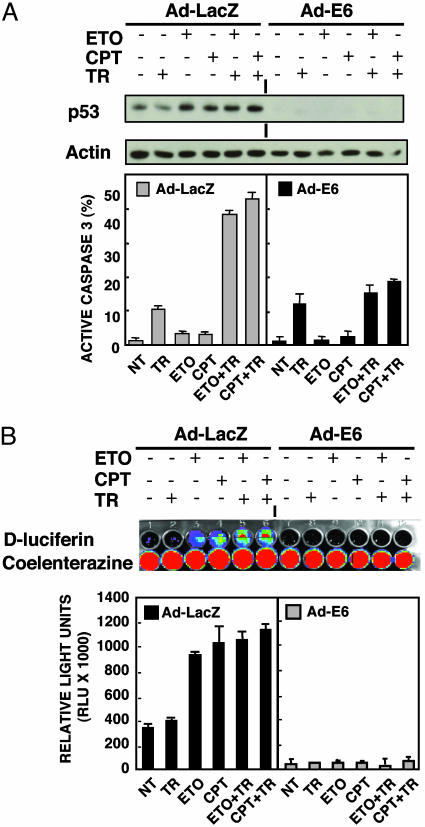
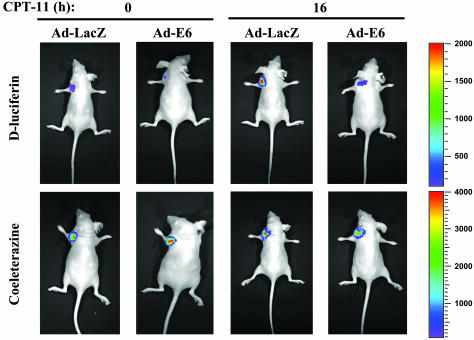
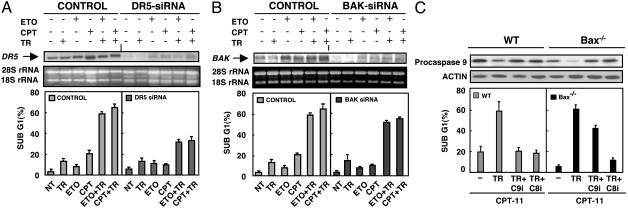
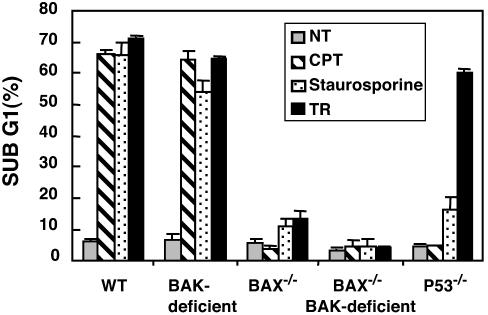
Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) exhibits specific tumoricidal activity and is under development for cancer therapy. Mismatch-repair-deficient colonic tumors evade TRAIL-induced apoptosis through mutational inactivation of Bax, but chemotherapeutics including Camptosar (CPT-11) restore TRAIL sensitivity. However, the signaling pathways in restoring TRAIL sensitivity remain to be elucidated. Here, we imaged p53 transcriptional activity in Bax-/- carcinomas by using bioluminescence, in vivo, and find that p53 is required for sensitization to TRAIL by CPT-11. Small interfering RNAs directed at proapoptotic p53 targets reveal TRAIL receptor KILLER/DR5 contributes significantly to TRAIL sensitization, whereas Bak plays a minor role. Caspase 8 inhibition protects both CPT-11 pretreated wild-type and Bax-/- HCT116 cells from TRAIL-induced apoptosis, whereas caspase 9 inhibition only rescued the wild-type HCT116 cells from death induced by TRAIL. The results suggest a conversion in the apoptotic mechanism in HCT116 colon carcinoma from a type II pathway involving Bax and the mitochondria to a type I pathway involving efficient extrinsic pathway caspase activation. In contrast to Bax-/- cells, Bak-deficient human cancers undergo apoptosis in response to TRAIL or CPT-11, implying that these proteins have nonoverlapping functions. Our studies elucidate a mechanism for restoration of TRAIL sensitivity in MMR-deficient Bax-/- human cancers through p53-dependent activation of KILLER/DR5 and reconstitution of a type I death pathway. Efforts to identify agents that up-regulate DR5 may be useful in cancer therapies restoring TRAIL sensitivity.
Figures
References
-
- Walczak, H., Miller, R. E., Ariail, K., Gliniak, B., Griffith, T. S., Kubin, M., Chin, W., Jones, J., Woodward, A., Le, T., et al. (1999) Nat. Med. 5, 157-163. - PubMed
-
- Takeda, K., Hayakawa, Y., Smyth, M. J., Kayagaki, N., Yamaguchi, N., Kakuta, S., Iwakura, Y., Yagita, H. & Okumura, K. (2001) Nat. Med. 7, 94-100. - PubMed
-
- Ashkenazi, A. & Dixit, V. M. (1998) Science 281, 1305-1308. - PubMed
-
- Wu, G. S., Burns, T. F., McDonald, E. R., III, Jiang, W., Meng, R., Krantz, I. D., Kao, G., Gan, D. D., Zhou, J. Y., Muschel, R., et al. (1997) Nat. Genet. 17, 141-143. - PubMed
-
- Ichikawa, K., Liu, W., Zhao, L., Wang, Z., Liu, D., Ohtsuka, T., Zhang, H., Mountz, J. D., Koopman, W. J., Kimberly, R. P. & Zhou, T. (2001) Nat. Med. 7, 954-960. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
