

Ribosome binding to the Oxa1 complex facilitates co-translational protein insertion in mitochondria
- PMID: 14657018
- PMCID: PMC291818
- DOI: 10.1093/emboj/cdg623
Ribosome binding to the Oxa1 complex facilitates co-translational protein insertion in mitochondria
Abstract
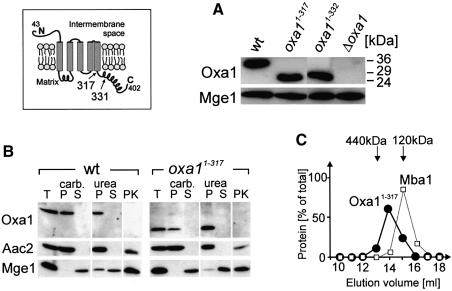
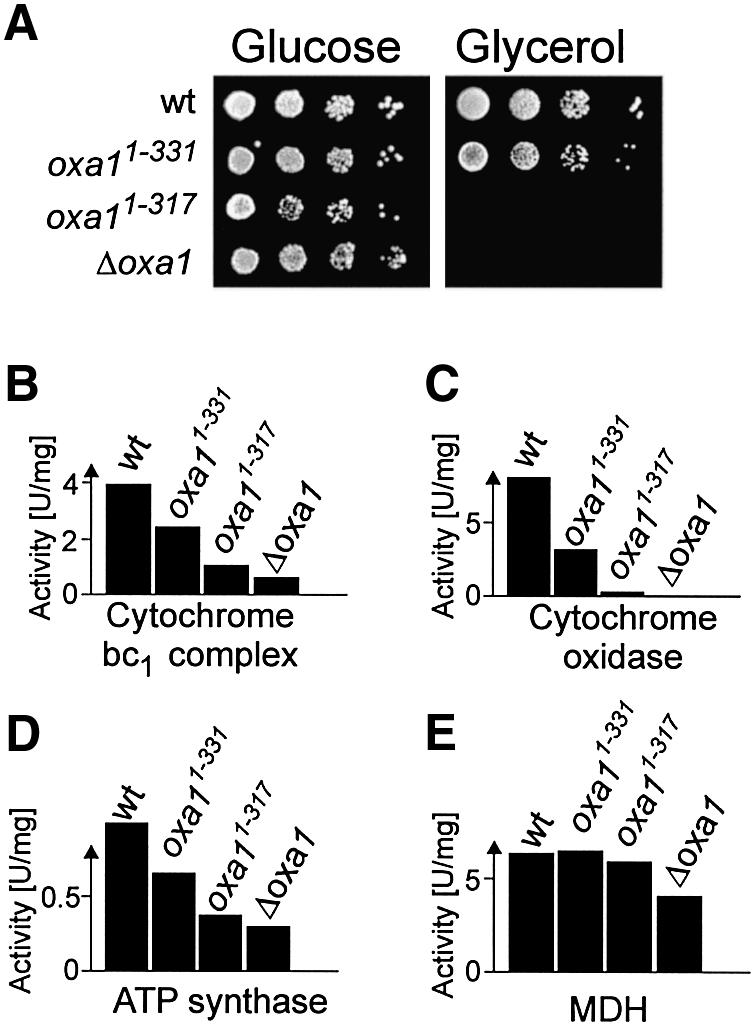
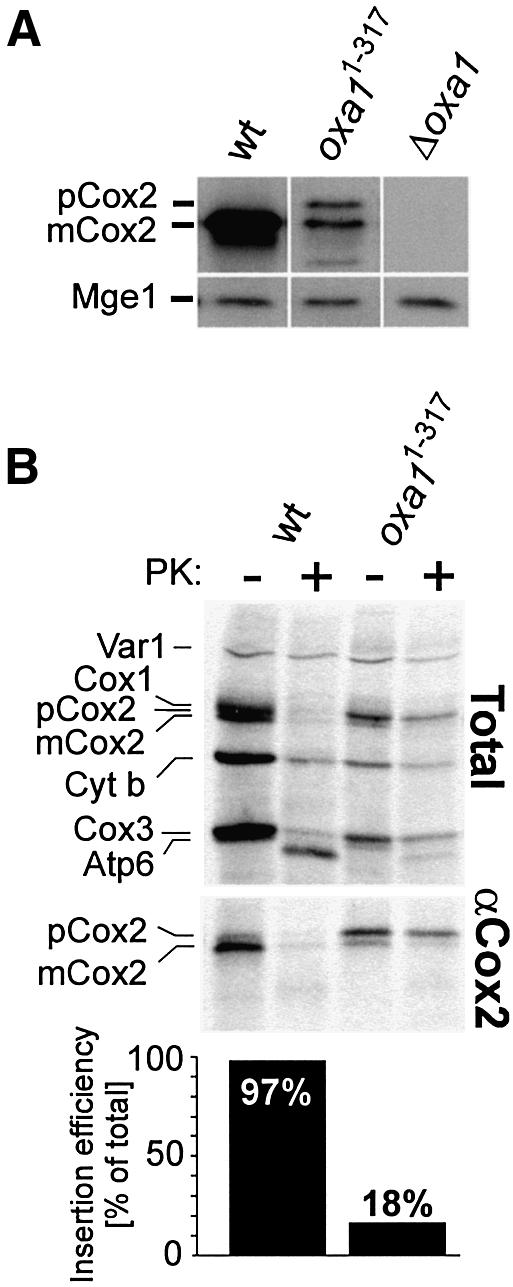
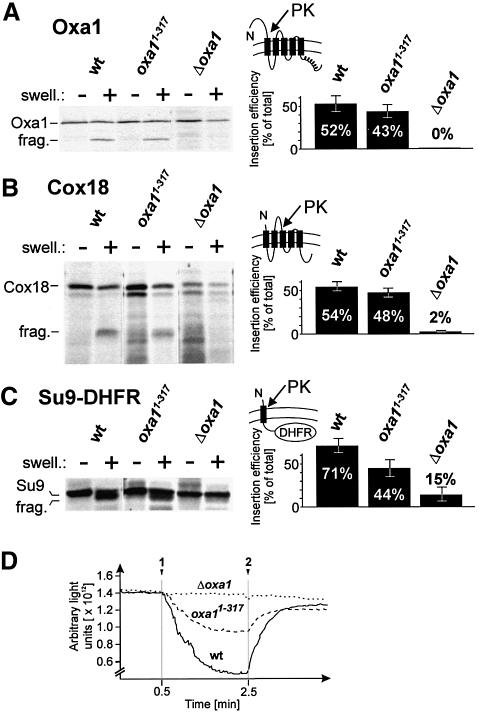
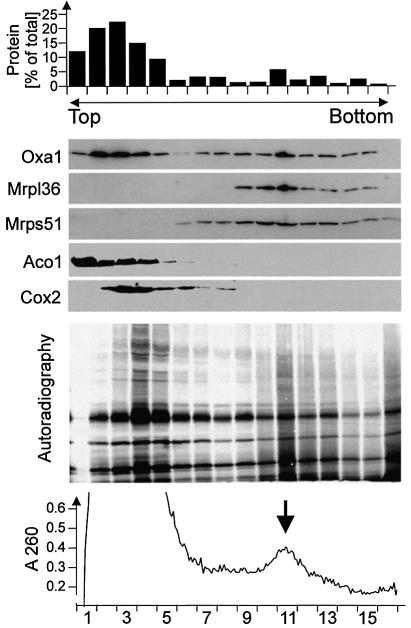
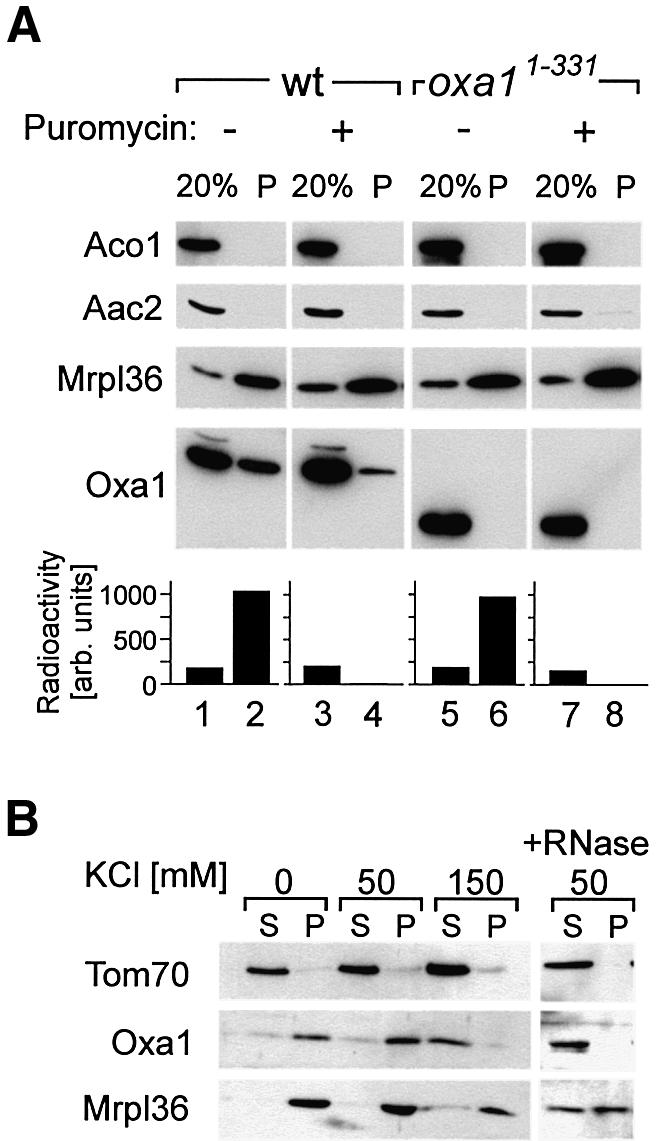
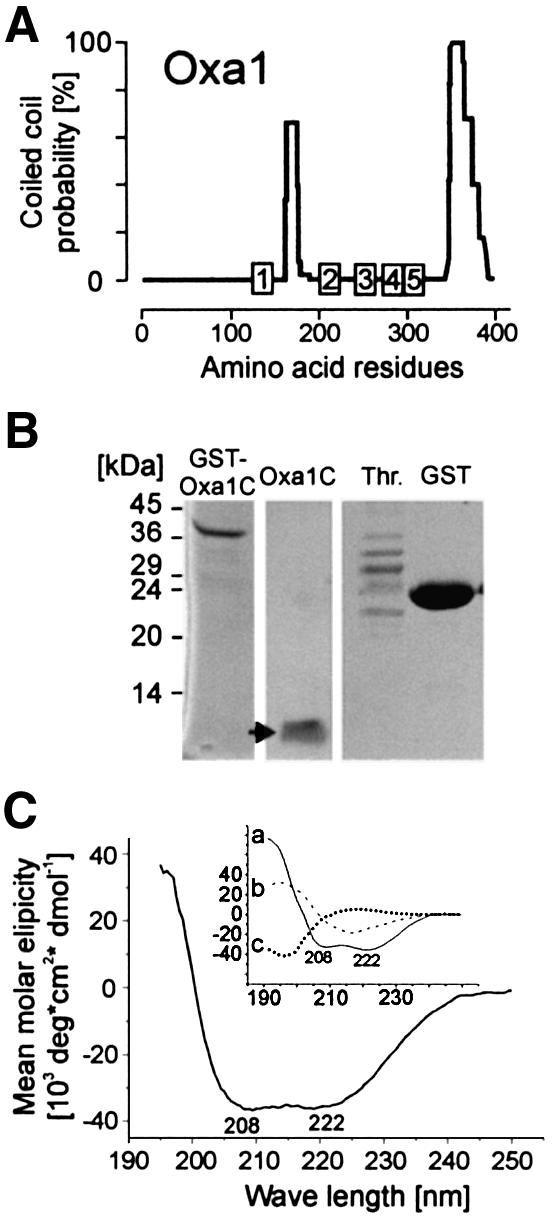
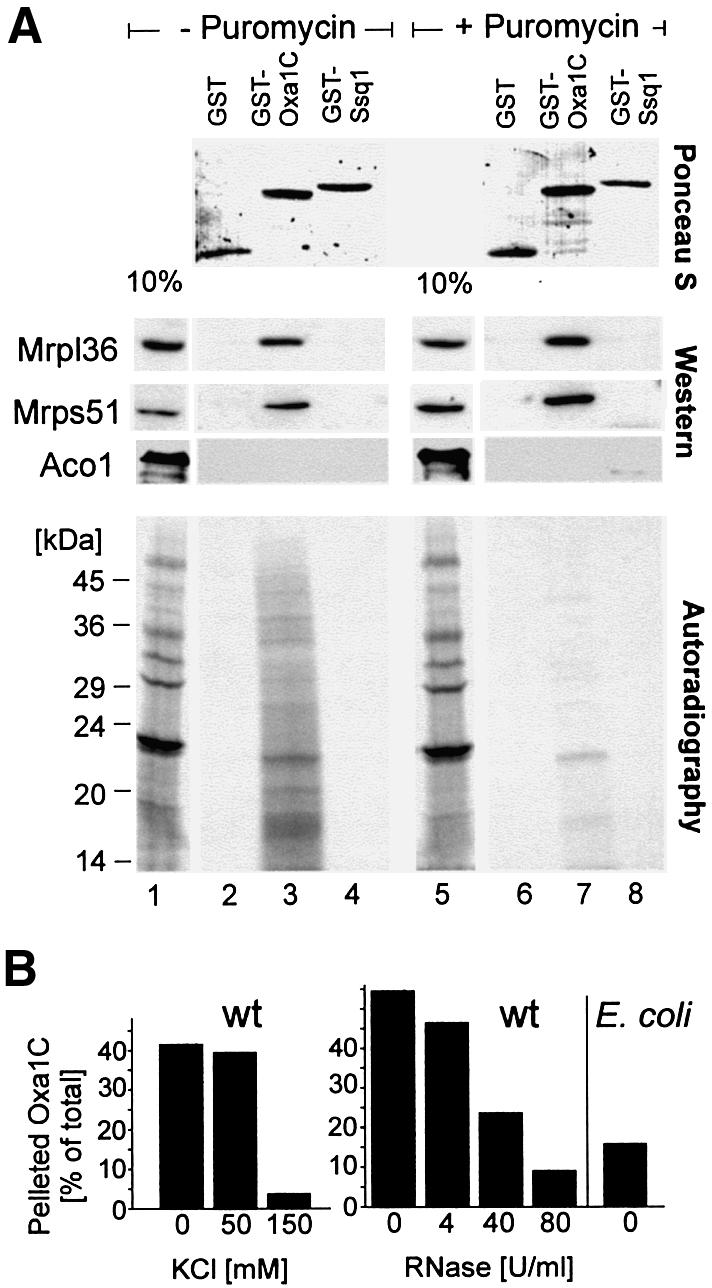
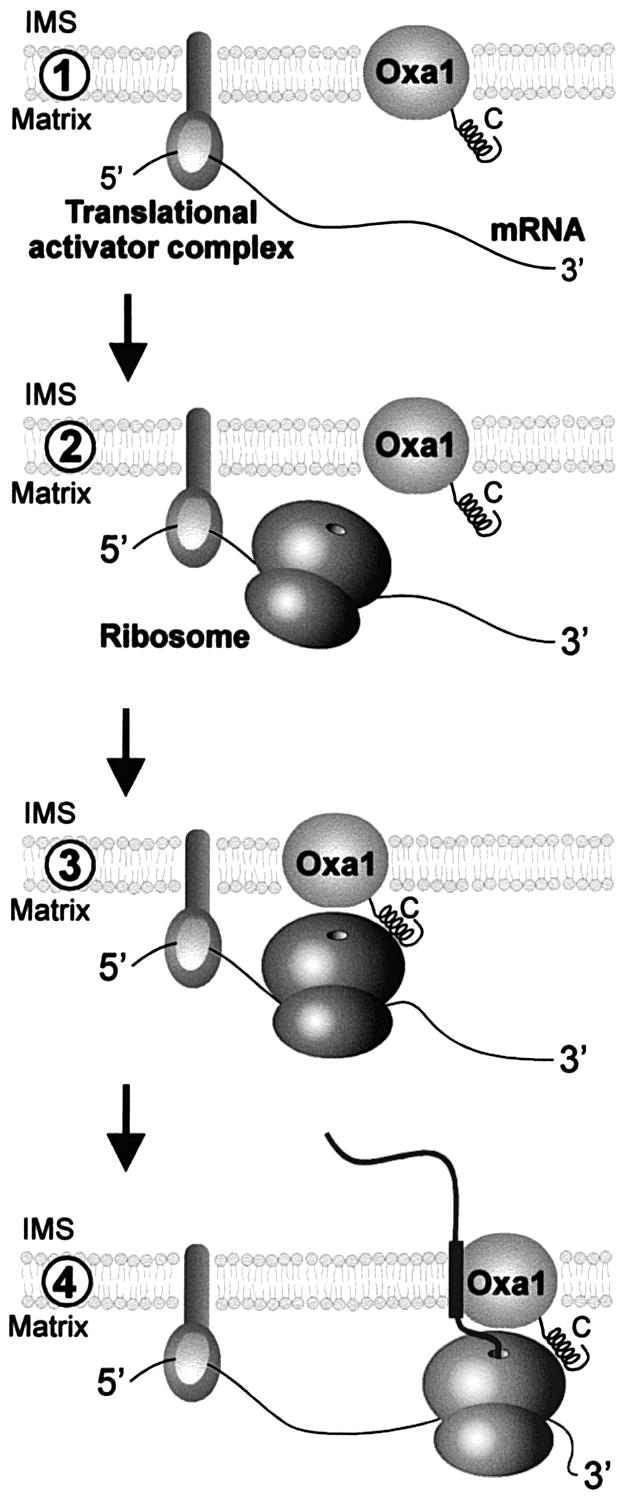
The Oxa1 translocase of the mitochondrial inner membrane facilitates the insertion of both mitochondrially and nuclear-encoded proteins from the matrix into the inner membrane. Most mitochondrially encoded proteins are hydrophobic membrane proteins which are integrated into the lipid bilayer during their synthesis on mitochondrial ribosomes. The molecular mechanism of this co-translational insertion process is unknown. Here we show that the matrix-exposed C-terminus of Oxa1 forms an alpha-helical domain that has the ability to bind to mitochondrial ribosomes. Deletion of this Oxa1 domain strongly diminished the efficiency of membrane insertion of subunit 2 of cytochrome oxidase, a mitochondrially encoded substrate of the Oxa1 translocase. This suggests that co-translational membrane insertion of mitochondrial translation products is facilitated by a physical interaction of translation complexes with the membrane-bound translocase.
Figures
References
-
- Bauer M., Behrens,M., Esser,K., Michaelis,G. and Pratje,E. (1994) PET1402, a nuclear gene required for proteolytic processing of cytochrome oxidase subunit 2 in yeast. Mol. Gen. Genet., 245, 272–278. - PubMed
-
- Beckmann R., Spahn,C.M., Eswar,N., Helmers,J., Penczek,P.A., Sali,A., Frank,J. and Blobel,G. (2001) Architecture of the protein-conducting channel associated with the translating 80S ribosome. Cell, 107, 361–372. - PubMed
-
- Bonnefoy N., Chalvet,F., Hamel,P., Slominski,P.P. and Dujardin,G. (1994) OXA1, a Saccharomyces cerevisiae nuclear gene whose sequence is conserved from prokaryotes to eukaryotes controls cytochrome oxidase biogenesis. J. Mol. Biol., 239, 201–212. - PubMed
-
- Borel A.C. and Simon,S.M. (1996) Biogenesis of polytopic membrane proteins: membrane segments assemble within translocation channels prior to membrane integration. Cell, 85, 379–389. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
