

Mouse Rev1 protein interacts with multiple DNA polymerases involved in translesion DNA synthesis
- PMID: 14657033
- PMCID: PMC291821
- DOI: 10.1093/emboj/cdg626
Mouse Rev1 protein interacts with multiple DNA polymerases involved in translesion DNA synthesis
Abstract
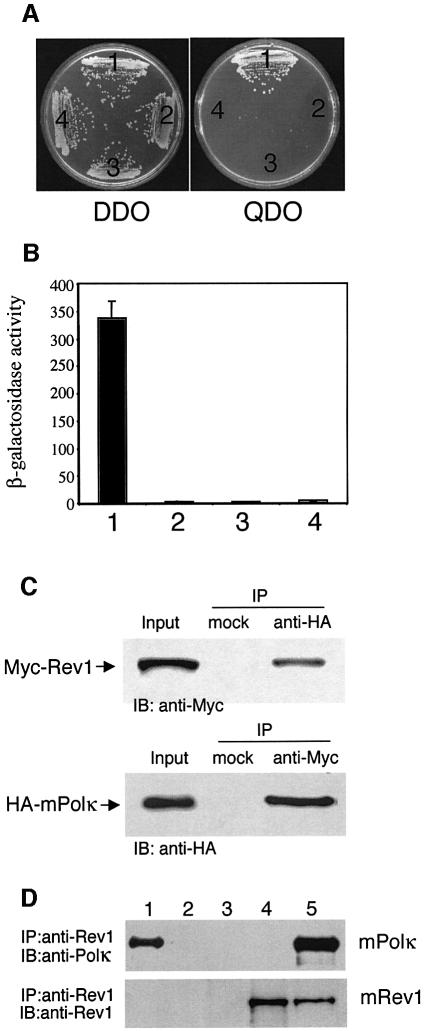
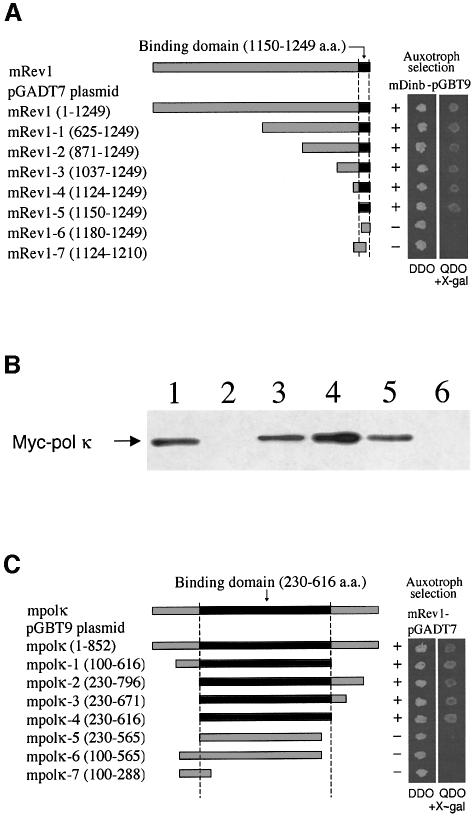
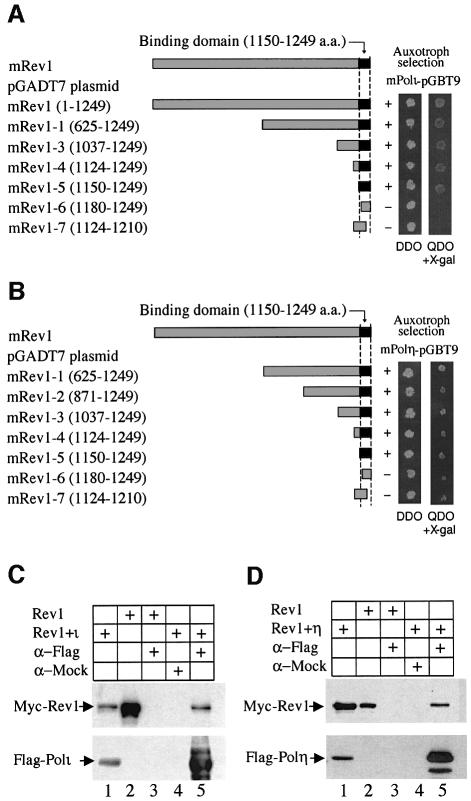
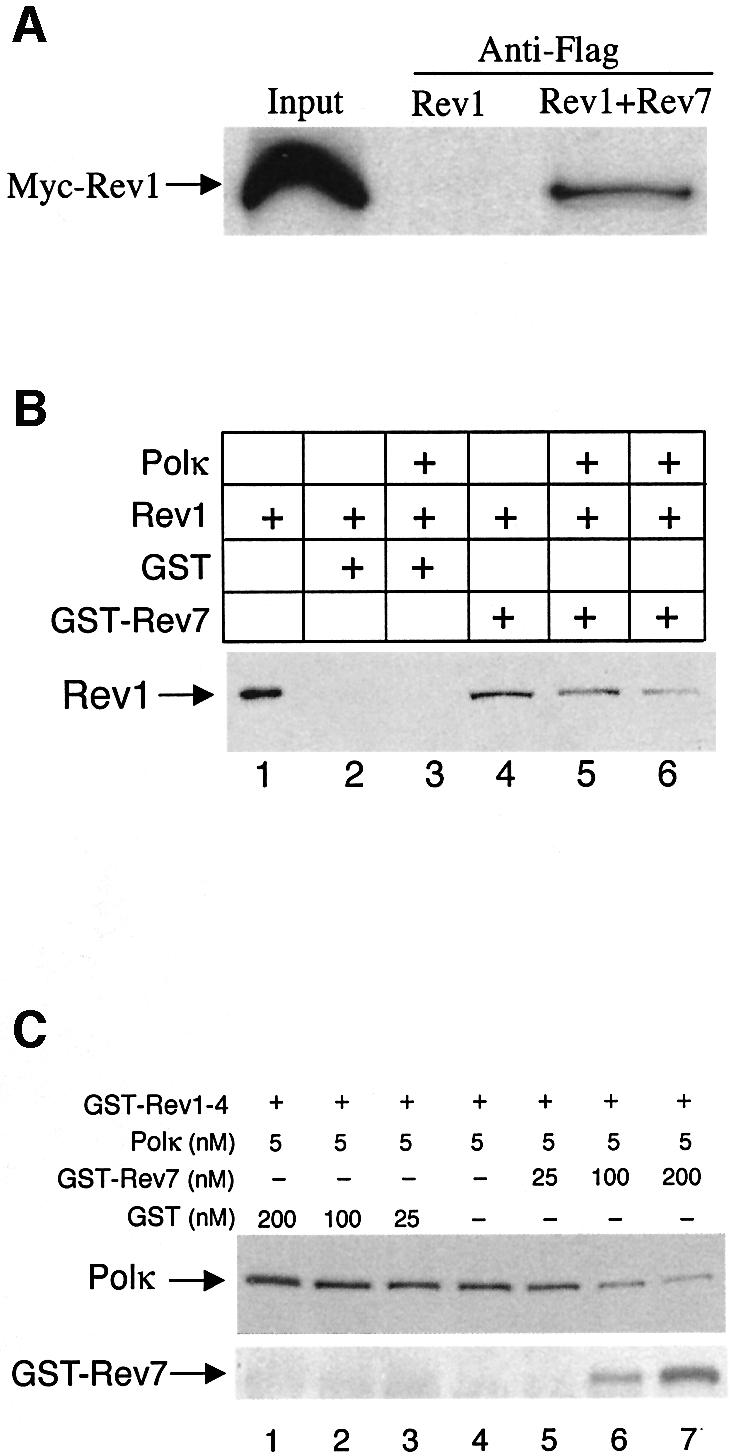
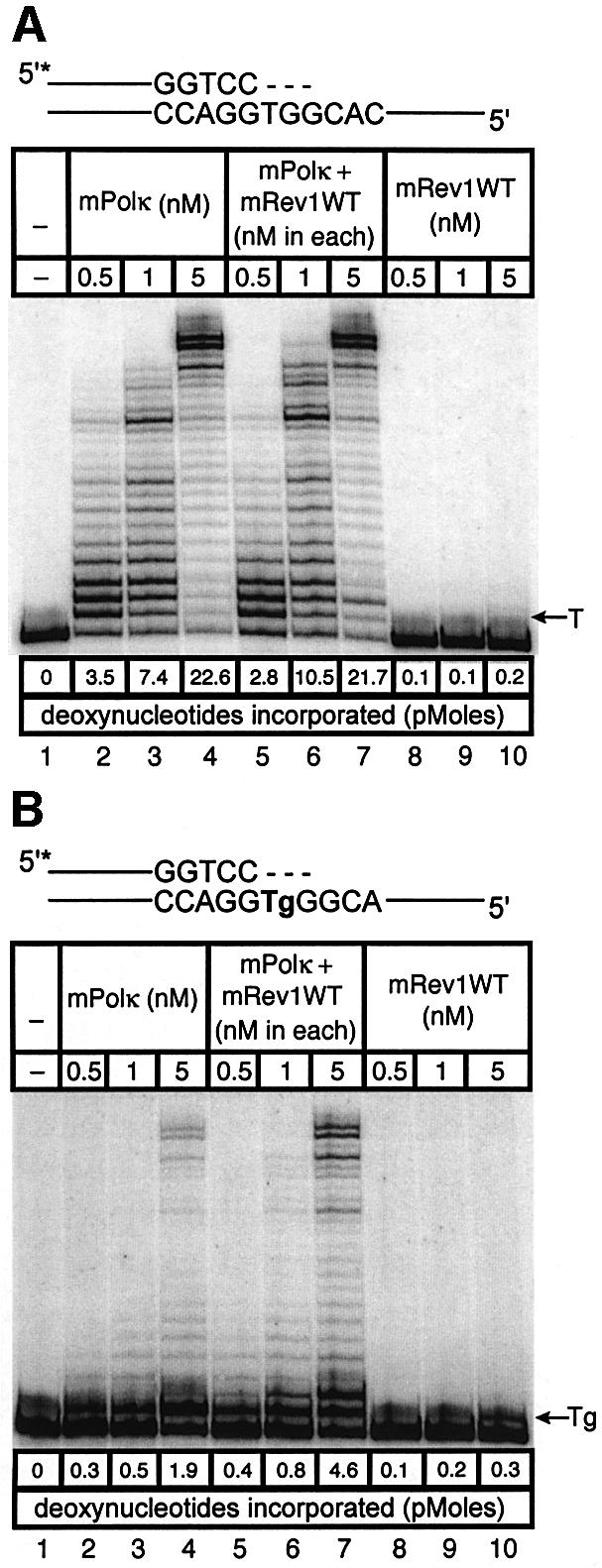
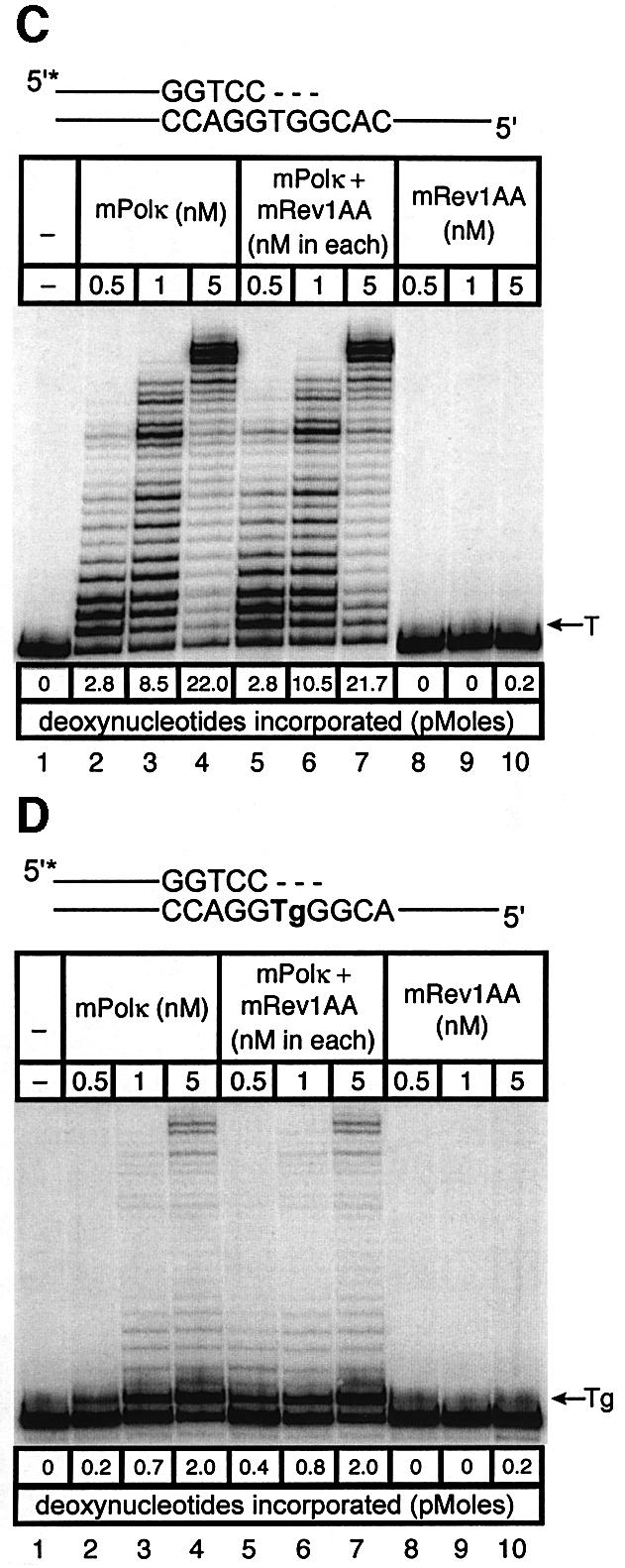
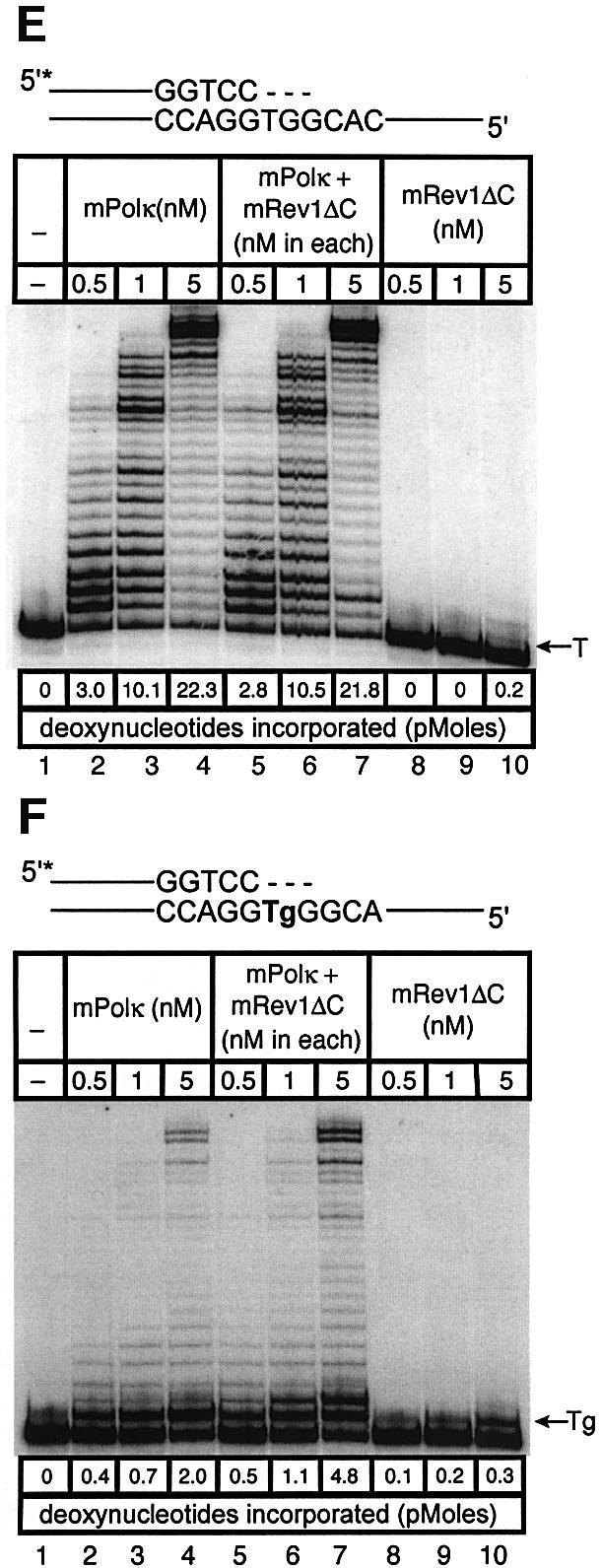
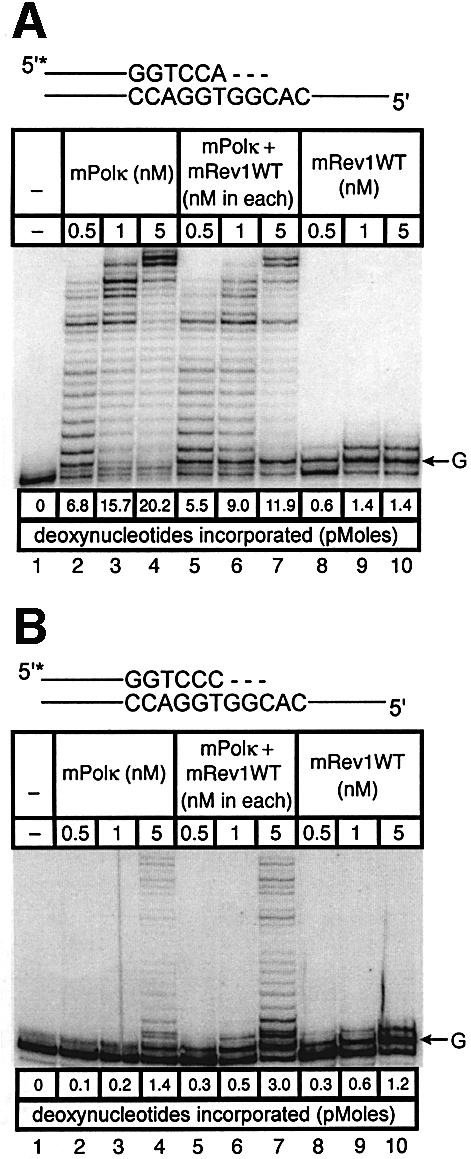
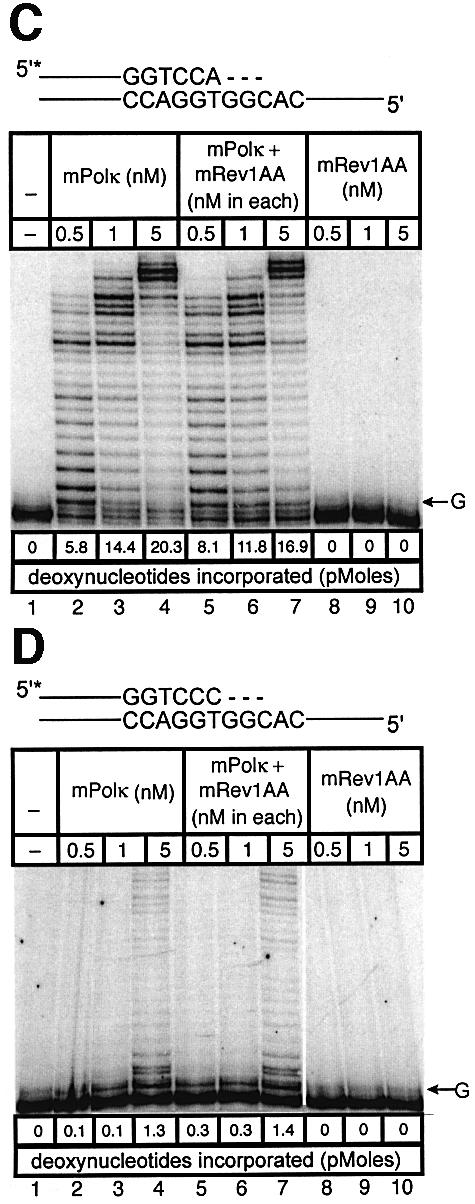
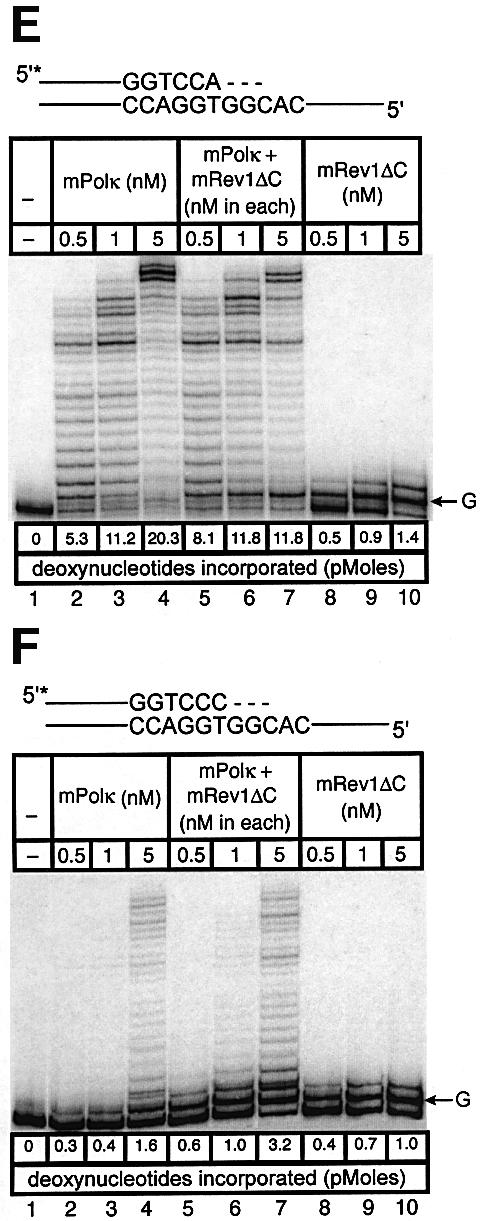
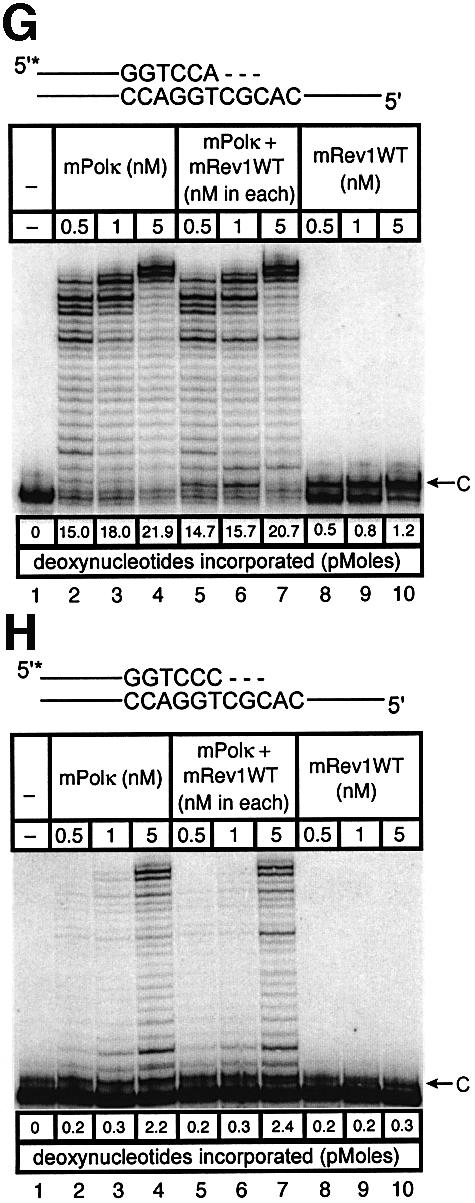
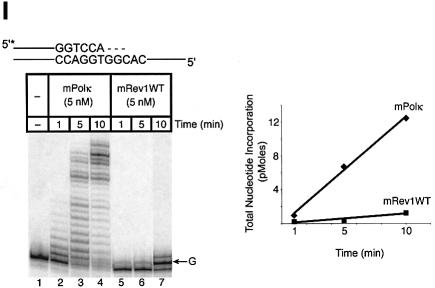
Pol kappa and Rev1 are members of the Y family of DNA polymerases involved in tolerance to DNA damage by replicative bypass [translesion DNA synthesis (TLS)]. We demonstrate that mouse Rev1 protein physically associates with Pol kappa. We show too that Rev1 interacts independently with Rev7 (a subunit of a TLS polymerase, Pol zeta) and with two other Y-family polymerases, Pol iota and Pol eta. Mouse Pol kappa, Rev7, Pol iota and Pol eta each bind to the same approximately 100 amino acid C-terminal region of Rev1. Furthermore, Rev7 competes directly with Pol kappa for binding to the Rev1 C-terminus. Notwithstanding the physical interaction between Rev1 and Pol kappa, the DNA polymerase activity of each measured by primer extension in vitro is unaffected by the complex, either when extending normal primer-termini, when bypassing a single thymine glycol lesion, or when extending certain mismatched primer termini. Our observations suggest that Rev1 plays a role(s) in mediating protein-protein interactions among DNA polymerases required for TLS. The precise function(s) of these interactions during TLS remains to be determined.
Figures
References
-
- Baynton K., Bresson-Roy,A. and Fuchs,R.P. (1999) Distinct roles for Rev1p and Rev7p during translesion synthesis in Saccharomyces cerevisiae. Mol. Microbiol., 34, 124–133. - PubMed
-
- Bridges B.A. and Woodgate,R. (1985) The two-step model of bacterial UV mutagenesis. Mutat. Res., 150, 133–139. - PubMed
-
- Fischhaber P.L., Gerlach,V.L., Feaver,W.J., Hatahet,Z., Wallace,S.S. and Friedberg,E.C. (2002) Human DNA polymerase κ bypasses and extends beyond thymine glycols during translesion synthesis in vitro, preferentially incorporating correct nucleotides. J. Biol. Chem., 277, 37604–37611. - PubMed
-
- Friedberg E.C., Wagner,R. and Radman,M. (2002) Specialized DNA polymerases, cellular survival and the genesis of mutations. Science, 296, 1627–1630. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
