

CD40-deficient, influenza-specific CD8 memory T cells develop and function normally in a CD40-sufficient environment
- PMID: 14657225
- PMCID: PMC2194135
- DOI: 10.1084/jem.20031440
CD40-deficient, influenza-specific CD8 memory T cells develop and function normally in a CD40-sufficient environment
Abstract
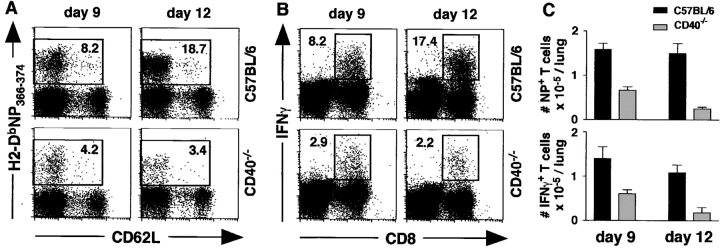
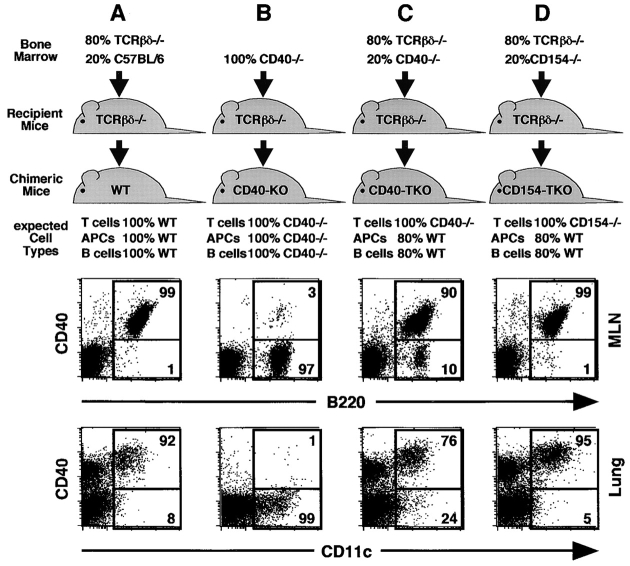
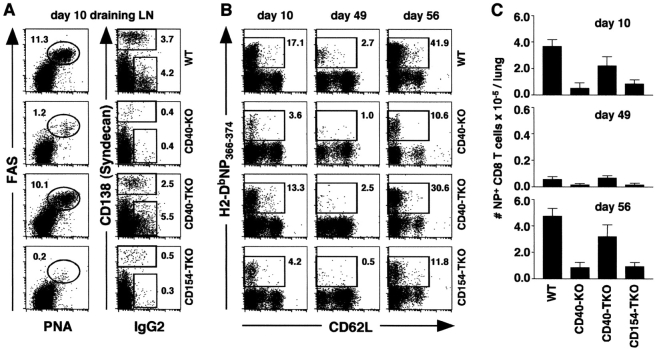
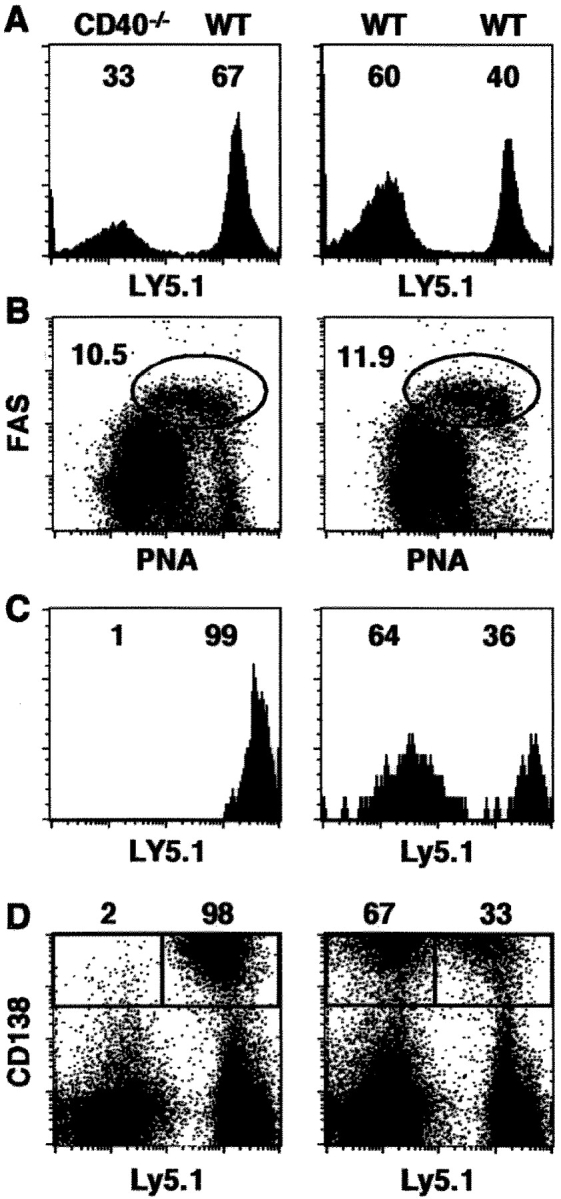
Two models have been proposed to explain the requirement for CD40 signaling in CD8 T cell responses. The first model suggests that CD4 T cells activate antigen-presenting cells (APCs) through CD40 signaling (APC licensing). In turn, licensed APCs are able to prime naive CD8 T cells. The second model suggests that CD154-expressing CD4 T cells activate CD40-bearing CD8 T cells directly. Although the requirement for CD40 in APC licensing can be bypassed by inflammatory responses to pathogens that activate APCs directly, the second model predicts that CD8 responses to all antigens will be dependent on CD40 signaling. Here we determined which model applies to CD8 responses to influenza. We demonstrate that optimal CD8 T cell responses to influenza are dependent on CD40 signaling, however both primary and secondary responses to influenza require CD40 expression on non-T cells. Furthermore, CD40-/- CD8 T cells proliferate and differentiate to the same extent as CD40+/+ CD8 T cells in response to influenza, as long as they have equal access to CD40+/+ APCs. Thus, CD4 T cells do not activate influenza-specific CD8 cells directly through CD40 signaling. Instead, these data support the classical model, in which CD4 T cells provide help to CD8 T cells indirectly by activating APCs through CD40.
Figures
Similar articles
-
Synergistic CD40 signaling on APCs and CD8 T cells drives efficient CD8 response and memory differentiation.J Leukoc Biol. 2012 Jun;91(6):859-69. doi: 10.1189/jlb.0611292. Epub 2012 Jan 11. J Leukoc Biol. 2012. PMID: 22241832
-
Influenza-induced, helper-independent CD8+ T cell responses use CD40 costimulation at the late phase of the primary response.J Leukoc Biol. 2013 Jan;93(1):145-54. doi: 10.1189/jlb.0612266. Epub 2012 Oct 29. J Leukoc Biol. 2013. PMID: 23108101
-
CD154+ CD4+ T-cell dependence for effective memory influenza virus-specific CD8+ T-cell responses.Immunol Cell Biol. 2014 Aug;92(7):605-11. doi: 10.1038/icb.2014.28. Epub 2014 Apr 29. Immunol Cell Biol. 2014. PMID: 24777309
-
Mini-review CD4 T cells are required for CD8 T cell memory generation.Eur J Immunol. 2003 Dec;33(12):3225-31. doi: 10.1002/eji.200324576. Eur J Immunol. 2003. PMID: 14635030 Review.
-
APC licensing and CD4+T cell help in liver-stage malaria.Front Microbiol. 2014 Nov 11;5:617. doi: 10.3389/fmicb.2014.00617. eCollection 2014. Front Microbiol. 2014. PMID: 25426113 Free PMC article. Review.
Cited by
-
Recall responses by helpless memory CD8+ T cells are restricted by the up-regulation of PD-1.J Immunol. 2009 Apr 1;182(7):4244-54. doi: 10.4049/jimmunol.0802041. J Immunol. 2009. PMID: 19299723 Free PMC article.
-
Bidirectional Communication Between the Innate and Adaptive Immune Systems.Annu Rev Immunol. 2025 Apr;43(1):489-514. doi: 10.1146/annurev-immunol-083122-040624. Annu Rev Immunol. 2025. PMID: 40279312 Free PMC article. Review.
-
CD40-CD40 ligand interactions promote trafficking of CD8+ T cells into the brain and protection against West Nile virus encephalitis.J Virol. 2007 Sep;81(18):9801-11. doi: 10.1128/JVI.00941-07. Epub 2007 Jul 11. J Virol. 2007. PMID: 17626103 Free PMC article.
-
Liposome-coupled peptides induce long-lived memory CD8 T cells without CD4 T cells.PLoS One. 2010 Nov 30;5(11):e15091. doi: 10.1371/journal.pone.0015091. PLoS One. 2010. PMID: 21264321 Free PMC article.
-
CD40-CD40L cross-talk drives fascin expression in dendritic cells for efficient antigen presentation to CD4+ T cells.Int Immunol. 2017 Mar 1;29(3):121-131. doi: 10.1093/intimm/dxx013. Int Immunol. 2017. PMID: 28369442 Free PMC article.
References
-
- Kawabe, T., T. Naka, K. Yoshida, T. Tanaka, H. Fujiwara, S. Suematsu, N. Yoshida, T. Kishimoto, and H. Kikutani. 1994. The immune responses in CD40-deficient mice: impaired immunoglobulin class switching and germinal center formation. Immunity. 1:167–178. - PubMed
-
- Bourgeois, C., B. Rocha, and C. Tanchot. 2002. A role for CD40 expression on CD8+ T cells in the generation of CD8+ T cell memory. Science. 297:2060–2063. - PubMed
-
- Ridge, J.P., F. Di Rosa, and P. Matzinger. 1998. A conditioned dendritic cell can be a temporal bridge between a CD4+ T-helper and a T-killer. Nature. 393:474–477. - PubMed
-
- Schoenberger, S.P., R.E.M. Toes, E.I.H. van der Voort, R. Offringa, and C.J.M. Melief. 1998. T-cell help for cytotoxic T lymphocytes is mediated by CD40-CD40L interactions. Nature. 393:480–483. - PubMed
-
- Bennett, S.R.M., F.R. Carbone, F. Karamalis, R.A. Flavell, J.F.A.P. Miller, and W.R. Heath. 1998. Help for cytotoxic-T-cell responses is mediated by CD40 signalling. Nature. 393:478–480. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
