

Direct sorting of the yeast uracil permease to the endosomal system is controlled by uracil binding and Rsp5p-dependent ubiquitylation
- PMID: 14657252
- PMCID: PMC329401
- DOI: 10.1091/mbc.e03-04-0202
Direct sorting of the yeast uracil permease to the endosomal system is controlled by uracil binding and Rsp5p-dependent ubiquitylation
Abstract
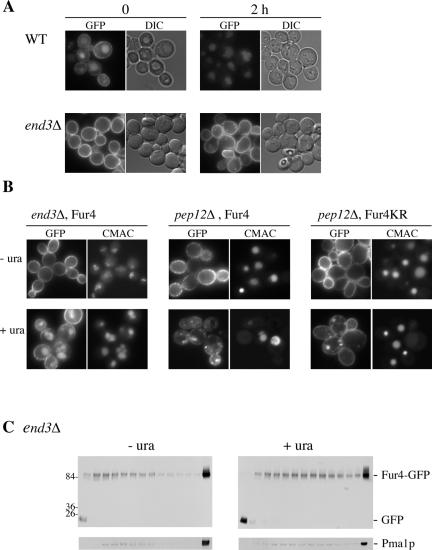
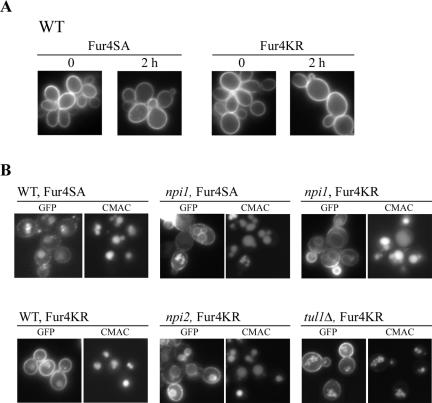
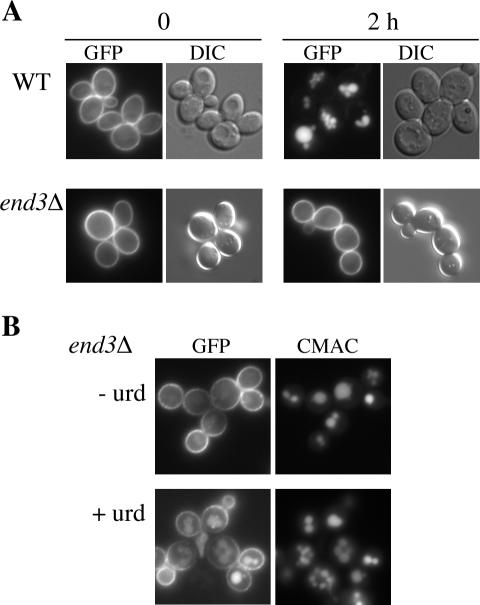
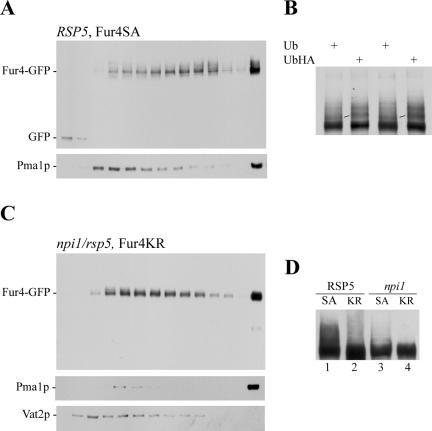
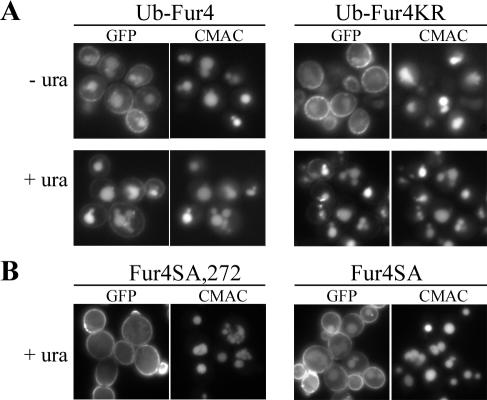
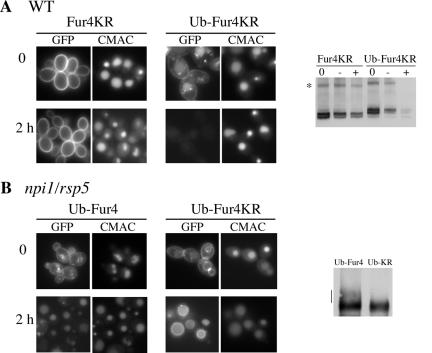
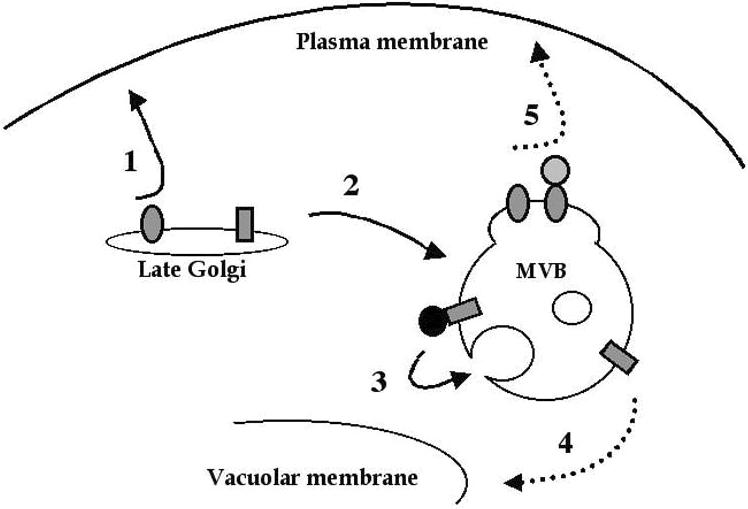
The yeast uracil permease, Fur4p, is downregulated by uracil, which is toxic to cells with high permease activity. Uracil promotes cell surface Rsp5p-dependent ubiquitylation of the permease, signaling its endocytosis and further vacuolar degradation. We show here that uracil also triggers the direct routing of its cognate permease from the Golgi apparatus to the endosomal system for degradation, without passage via the plasma membrane. This early sorting was not observed for a variant permease with a much lower affinity for uracil, suggesting that uracil binding is the signal for the diverted pathway. The FUI1-encoded uridine permease is similarly sorted for early vacuolar degradation in cells exposed to a toxic level of uridine uptake. Membrane proteins destined for vacuolar degradation require sorting at the endosome level to the intraluminal vesicles of the multivesicular bodies. In cells with low levels of Rsp5p, Fur4p can be still diverted from the Golgi apparatus but does not reach the vacuolar lumen, being instead missorted to the vacuolar membrane. Correct luminal delivery is restored by the biosynthetic addition of a single ubiquitin, suggesting that the ubiquitylation of Fur4p serves as a specific signal for sorting to the luminal vesicles of the multivesicular bodies. A fused ubiquitin is also able to sort some Fur4p from the Golgi to the degradative pathway in the absence of added uracil but the low efficiency of this sorting indicates that ubiquitin does not itself act as a dominant signal for Golgi-to-endosome trafficking. Our results are consistent with a model in which the binding of intracellular uracil to the permease signals its sorting from the Golgi apparatus and subsequent ubiquitylation ensures its delivery to the vacuolar lumen.
Figures
References
-
- Arvan, P., Zhao, X., Ramos-Castaneda, J., and Chang, A. (2002). Secretory pathway quality control operating in Golgi, plasmalemmal, and endosomal systems. Traffic 3, 771-80. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
