

Light- and pH-dependent structural changes in the PsbS subunit of photosystem II
- PMID: 14657329
- PMCID: PMC299978
- DOI: 10.1073/pnas.2533072100
Light- and pH-dependent structural changes in the PsbS subunit of photosystem II
Abstract
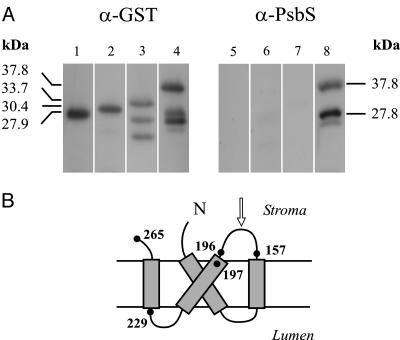
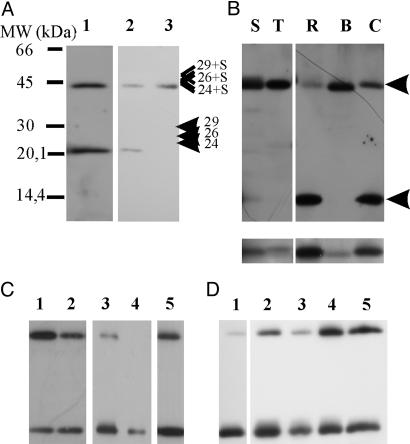
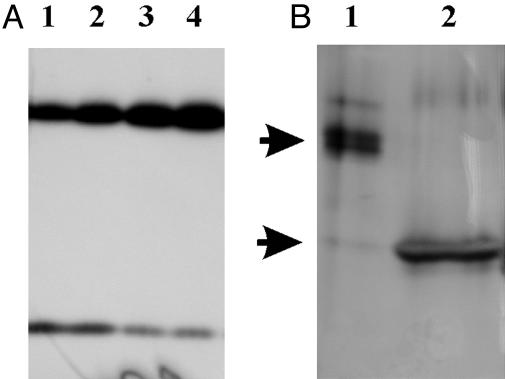
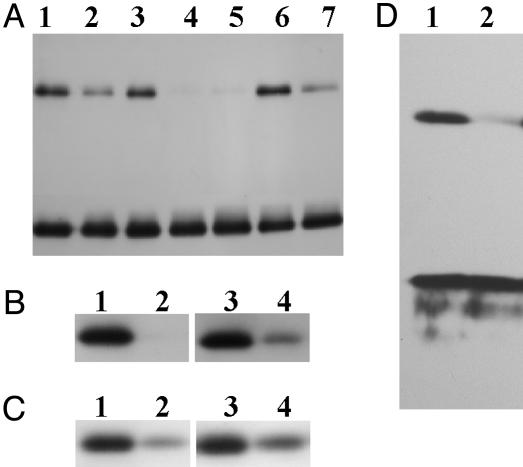
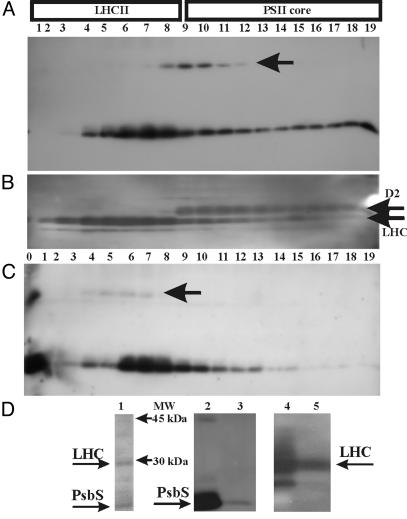
In higher plants, the PsbS subunit of photosystem II (PSII) plays a crucial role in pH- and xanthophyll-dependent nonphotochemical quenching of excess absorbed light energy, thus contributing to the defense mechanism against photoinhibition. We determined the amino acid sequence of Zea mays PsbS and produced an antibody that recognizes with high specificity a region of the protein located in the stroma-exposed loop between the second and third putative helices. By means of this antiserum, the thylakoid membranes of various higher plant species revealed the presence of a 42-kDa protein band, indicating the formation of a dimer of the 21-kDa PsbS protein. Crosslinking experiments and immunoblotting with other antisera seem to exclude the formation of a heterodimer with other PSII protein components. The PsbS monomer/dimer ratio in isolated thylakoid membranes was found to vary with luminal pH in a reversible manner, the monomer being the prevalent form at acidic and the dimer at alkaline pH. In intact chloroplasts and whole plants, dimer-to-monomer conversion is reversibly induced by light, known to cause luminal acidification. Sucrose-gradient centrifugation revealed a prevalent association of the PsbS monomer and dimer with light-harvesting complex and PSII core complexes, respectively. The finding of the existence of a light-induced change in the quaternary structure of the PsbS subunit may contribute to understanding the mechanism of PsbS action during nonphotochemical quenching.
Figures
References
-
- Barber, J. & Andersson, B. (1992) Trends Biochem. Sci. 17, 61-66. - PubMed
-
- Aro, E. M., Virgin, I. & Andersson, B. (1993) Biochim. Biophys. Acta 1143, 113-134. - PubMed
-
- Horton, P., Ruban, A. V. & Walters, R. G. (1996) Annu. Rev. Plant Physiol. Mol. Biol. 47, 655-684. - PubMed
-
- Niyogi, K. K. (1999) Annu. Rev. Plant Physiol. Mol. Biol. 50, 333-359. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
