

A methyl viologen-resistant mutant of Arabidopsis, which is allelic to ozone-sensitive rcd1, is tolerant to supplemental ultraviolet-B irradiation
- PMID: 14657410
- PMCID: PMC316307
- DOI: 10.1104/pp.103.033480
A methyl viologen-resistant mutant of Arabidopsis, which is allelic to ozone-sensitive rcd1, is tolerant to supplemental ultraviolet-B irradiation
Abstract
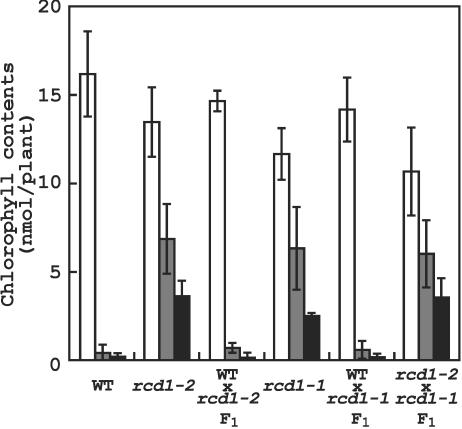
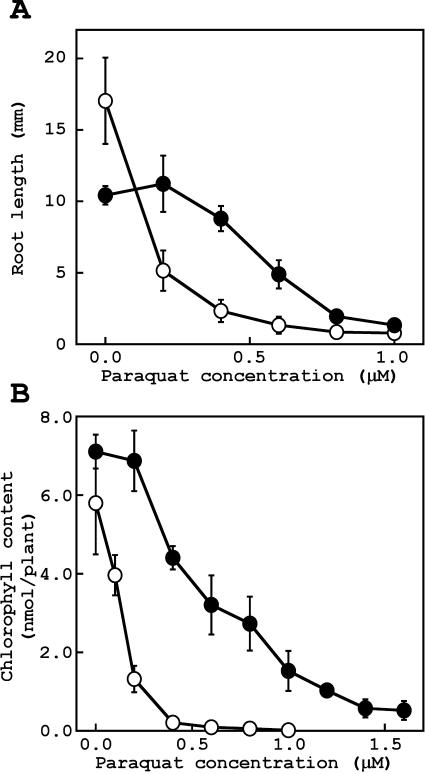
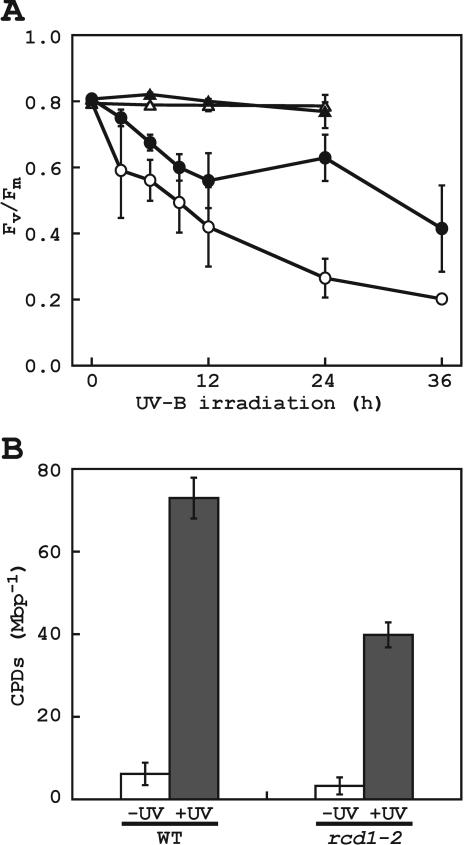
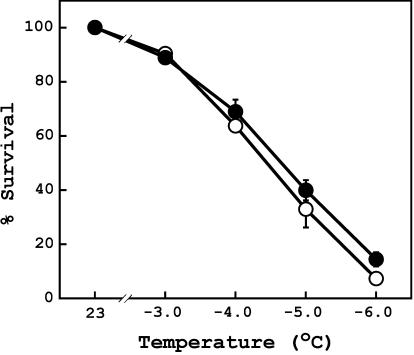
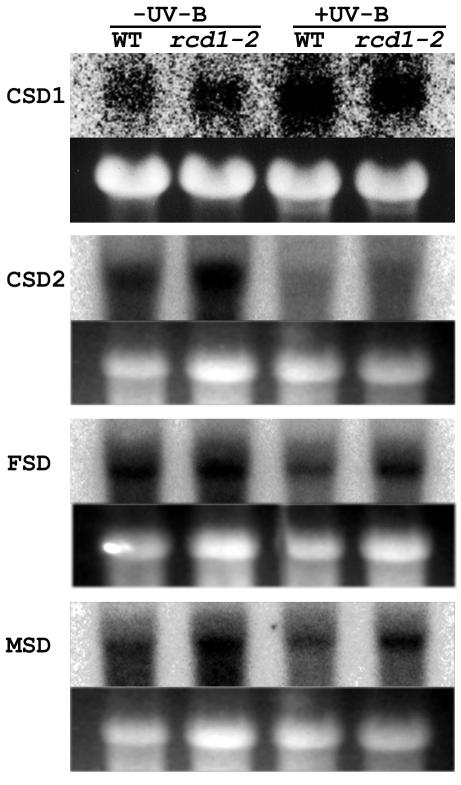
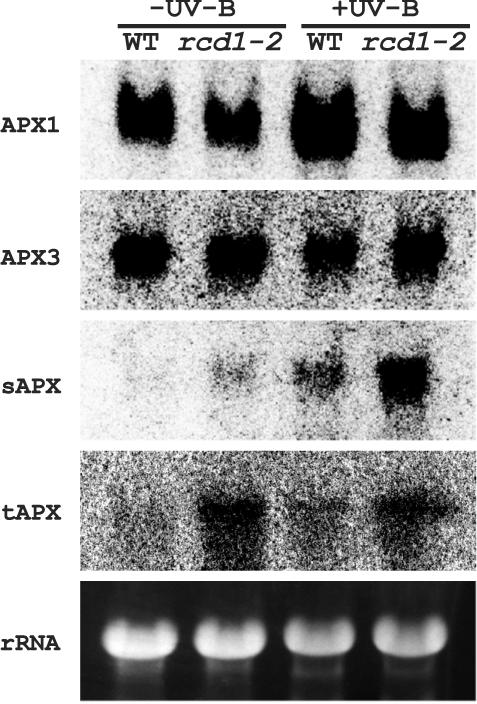
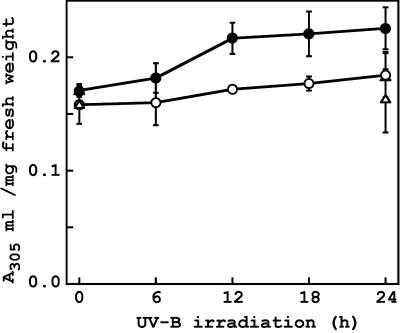
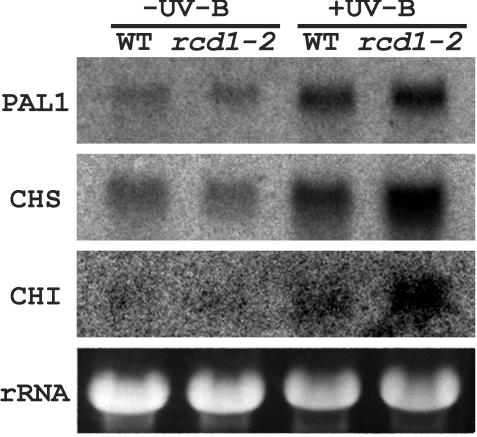
To better understand the role of active oxygen species (AOS) in acquired resistance to increased levels of ultraviolet (UV)-B irradiation in plants, we isolated an Arabidopsis mutant that is resistant to methyl viologen, and its sensitivity to UV-B was investigated. A complementation test revealed that the obtained mutant was allelic to the ozone-sensitive radical-induced cell death1-1 (rcd1-1). Therefore, this mutant was named rcd1-2. rcd1-2 was recessive and nearly 4-fold more resistant to methyl viologen than wild type. It exhibited a higher tolerance to short-term UV-B supplementation treatments than the wild type: UV-B-induced formation of cyclobutane pyrimidine dimers was reduced by one-half after 24 h of exposure; the decrease in quantum yield of photosystem II was also diminished by 40% after 12 h of treatment. Furthermore, rcd1-2 was tolerant to freezing. Steady-state mRNA levels of plastidic Cu/Zn superoxide dismutase and stromal ascorbate peroxidase were higher in rcd1-2 than in wild type, and the mRNA level of the latter enzyme was enhanced by UV-B exposure more effectively in rcd1-2. UV-B-absorbing compounds were more accumulated in rcd1-2 than in wild type after UV-B exposure for 24 h. These findings suggest that rcd1-2 methyl viologen resistance is due to the enhanced activities of the AOS-scavenging enzymes in chloroplasts and that the acquired tolerance to the short-term UV-B exposure results from a higher accumulation of sunscreen pigments. rcd1 appears to be a mutant that constitutively shows stress responses, leading to accumulation of more pigments and AOS-scavenging enzymes without any stresses.
Figures


References
-
- Abarca D, Roldán M, Martín M, Sabater B (2001) Arabidopsis thaliana ecotype Cvi shows an increased tolerance to photo-oxidative stress and contains a new chloroplastic copper/zinc superoxide dismutase isoenzyme. J Exp Bot 360: 1417-1425 - PubMed
-
- Asada K (1999) The water-water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Annu Rev Plant Physiol Plant Mol Biol 50: 601-639 - PubMed
-
- Britt AB, Chen JJ, Wykoff D, Mitchell D (1993) A UV-sensitive mutant of Arabidopsis defective in the repair of pyrimidine-pyrimidinone (6-4) dimers. Science 261: 1571-1574 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
