

No tail co-operates with non-canonical Wnt signaling to regulate posterior body morphogenesis in zebrafish
- PMID: 14660439
- PMCID: PMC5045862
- DOI: 10.1242/dev.00915
No tail co-operates with non-canonical Wnt signaling to regulate posterior body morphogenesis in zebrafish
Abstract
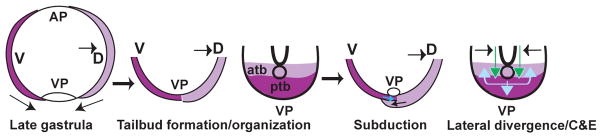
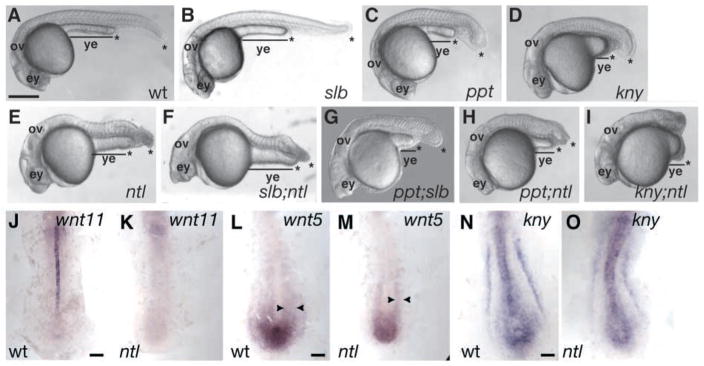
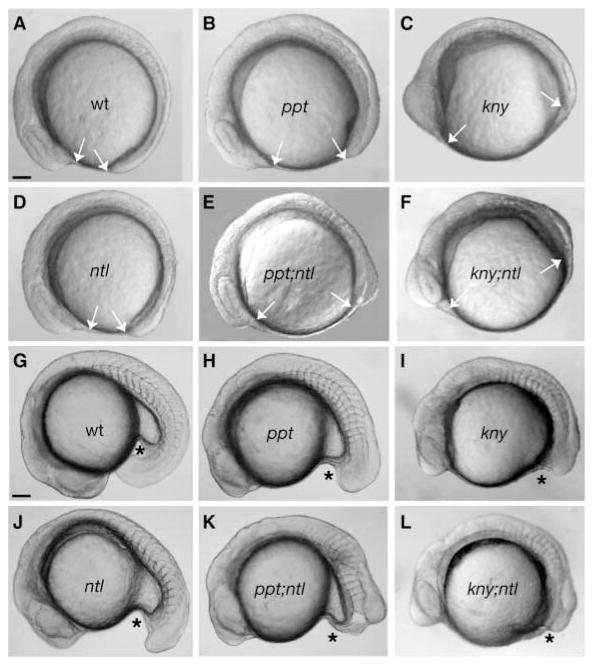
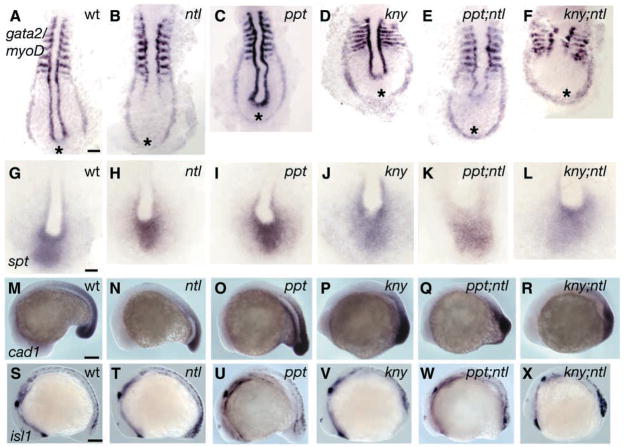
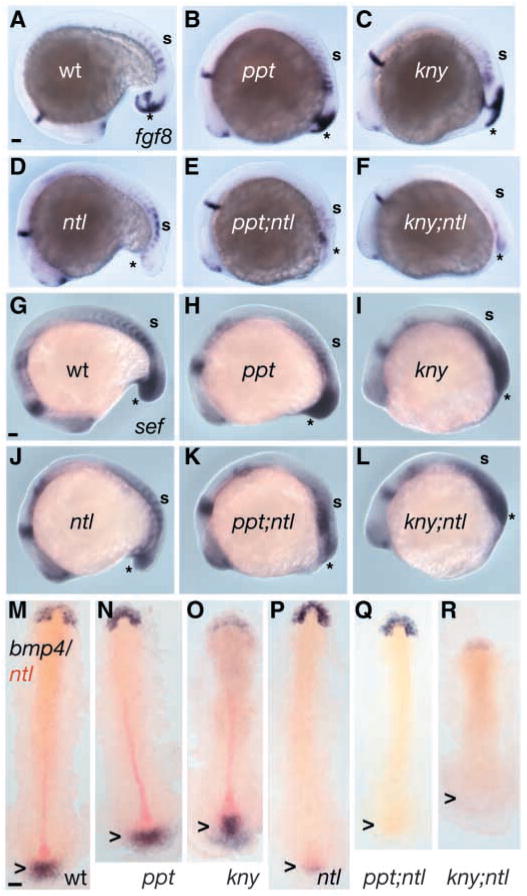
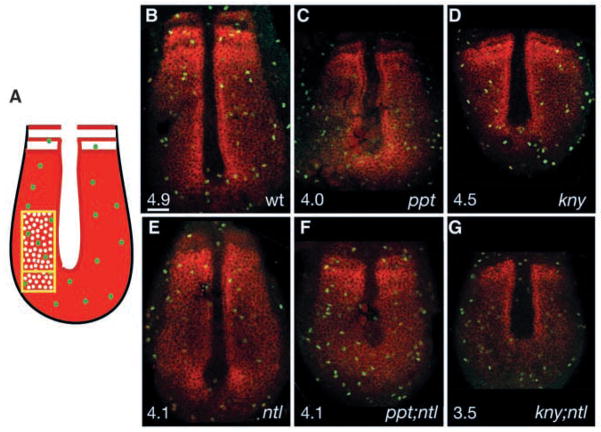
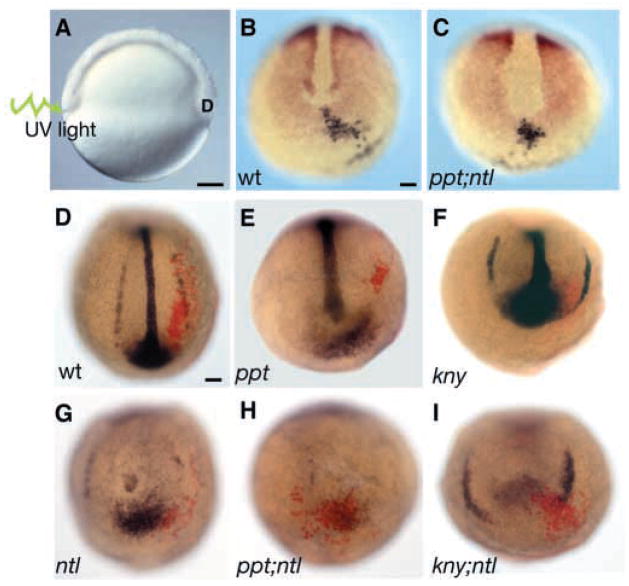
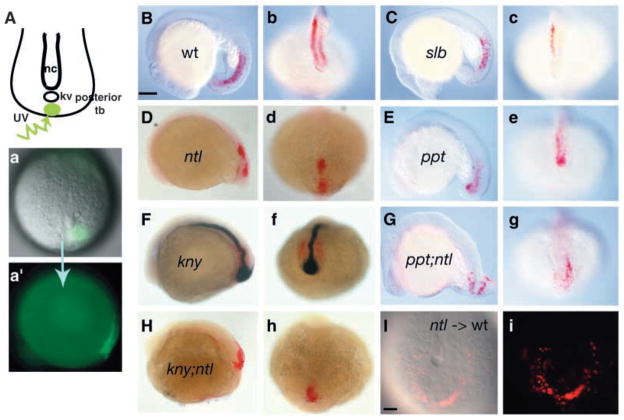
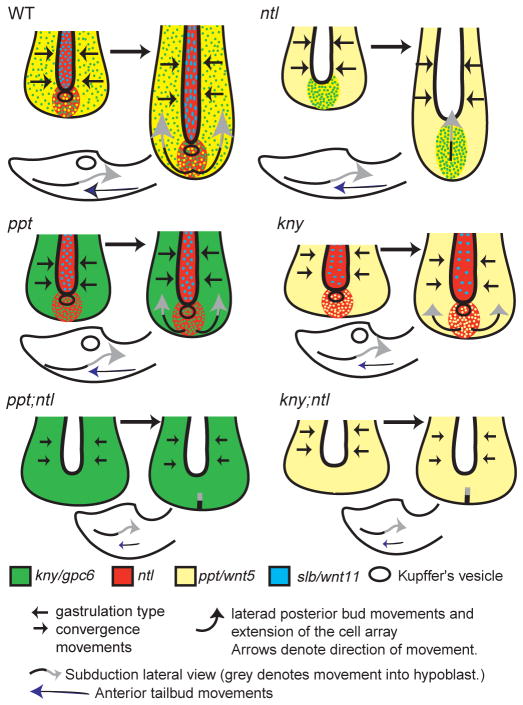
The vertebrate posterior body is formed by a combination of the gastrulation movements that shape the head and anterior trunk and posterior specific cell behaviors. Here, we investigated whether genes that regulate cell movements during gastrulation [no tail (ntl)/brachyury, knypek (kny) and pipetail (ppt)/wnt5] interact to regulate posterior body morphogenesis. Both kny;ntl and ppt;ntl double mutant embryos exhibit synergistic trunk and tail shortening by early segmentation. Gene expression analysis in the compound mutants indicates that anteroposterior germ-layer patterning is largely normal and that the tail elongation defects are not due to failure to specify or maintain posterior tissues. Moreover, ntl interacts with ppt and kny to synergistically regulate the posterior expression of the gene encoding bone morphogenetic protein 4 (bmp4) but not of other known T-box genes, fibroblast growth factor genes or caudal genes. Examination of mitotic and apoptotic cells indicates that impaired tail elongation is not simply due to decreased cell proliferation or increased cell death. Cell tracing in ppt;ntl and kny;ntl mutants demonstrates that the ventral derived posterior tailbud progenitors move into the tailbud. However, gastrulation-like convergence and extension movements and cell movements within the posterior tailbud are impaired. Furthermore, subduction movements of cells into the mesendoderm are reduced in kny;ntl and ppt;ntl mutants. We propose that Ntl and the non-canonical Wnt pathway components Ppt and Kny function in parallel, partially redundant pathways to regulate posterior body development. Our work initiates the genetic dissection of posterior body morphogenesis and links genes to specific tail-forming movements. Moreover, we provide genetic evidence for the notion that tail development entails a continuation of mechanisms regulating gastrulation together with mechanisms unique to the posterior body.
Figures
Similar articles
-
The role of Ppt/Wnt5 in regulating cell shape and movement during zebrafish gastrulation.Mech Dev. 2003 Apr;120(4):467-76. doi: 10.1016/s0925-4773(03)00004-2. Mech Dev. 2003. PMID: 12676324
-
Fyn/Yes and non-canonical Wnt signalling converge on RhoA in vertebrate gastrulation cell movements.EMBO Rep. 2005 May;6(5):426-31. doi: 10.1038/sj.embor.7400386. EMBO Rep. 2005. PMID: 15815683 Free PMC article.
-
Bmp inhibition is necessary for post-gastrulation patterning and morphogenesis of the zebrafish tailbud.Dev Biol. 2009 May 1;329(1):55-63. doi: 10.1016/j.ydbio.2009.02.016. Epub 2009 Feb 21. Dev Biol. 2009. PMID: 19236859 Free PMC article.
-
Heads or tails? Amphioxus and the evolution of anterior-posterior patterning in deuterostomes.Dev Biol. 2002 Jan 15;241(2):209-28. doi: 10.1006/dbio.2001.0503. Dev Biol. 2002. PMID: 11784106 Review.
-
Setting up for gastrulation in zebrafish.Curr Top Dev Biol. 2020;136:33-83. doi: 10.1016/bs.ctdb.2019.08.002. Epub 2019 Sep 3. Curr Top Dev Biol. 2020. PMID: 31959294 Review.
Cited by
-
Exploring the Effects of Different Types of Surfactants on Zebrafish Embryos and Larvae.Sci Rep. 2015 Jun 8;5:10107. doi: 10.1038/srep10107. Sci Rep. 2015. PMID: 26053337 Free PMC article.
-
The differentiation and movement of presomitic mesoderm progenitor cells are controlled by Mesogenin 1.Development. 2012 Dec;139(24):4656-65. doi: 10.1242/dev.078923. Development. 2012. PMID: 23172917 Free PMC article.
-
Embryonic vascular disruption adverse outcomes: Linking high throughput signaling signatures with functional consequences.Reprod Toxicol. 2017 Jun;70:82-96. doi: 10.1016/j.reprotox.2017.05.005. Epub 2017 May 17. Reprod Toxicol. 2017. PMID: 28527947 Free PMC article.
-
Mechanical control of tissue shape and morphogenetic flows during vertebrate body axis elongation.Sci Rep. 2021 Apr 21;11(1):8591. doi: 10.1038/s41598-021-87672-3. Sci Rep. 2021. PMID: 33883563 Free PMC article.
-
BMP and non-canonical Wnt signaling are required for inhibition of secondary tail formation in zebrafish.Development. 2011 Jun;138(12):2601-11. doi: 10.1242/dev.058404. Development. 2011. PMID: 21610036 Free PMC article.
References
-
- Agathon A, Thisse C, Thisse B. The molecular nature of the zebrafish tail organizer. Nature, Lond. 2003;424:448–452. - PubMed
-
- Ajiro K, Yoda K, Utsumi K, Nishikawa Y. Alteration of cell cycle-dependent histone phosphorylations by okadaic acid. Induction of mitosis-specific H3 phosphorylation and chromatin condensation in mammalian interphase cells. J Biol Chem. 1996;271:13197–13201. - PubMed
-
- Amacher SL, Draper BW, Summers BR, Kimmel CB. The zebrafish T-box genes no tail and spadetail are required for development of trunk and tail mesoderm and medial floor plate. Development. 2002;129:3311–3323. - PubMed
-
- Amaya E, Musci TJ, Kirschner MW. Expression of a dominant negative mutant of the FGF receptor disrupts mesoderm formation in Xenopus embryos. Cell. 1991;66:257–270. - PubMed
-
- Amaya E, Stein PA, Musci TJ, Kirschner MW. FGF signalling in the early specification of mesoderm in Xenopus. Development. 1993;118:477–487. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
