

Membrane tube formation from giant vesicles by dynamic association of motor proteins
- PMID: 14663143
- PMCID: PMC307611
- DOI: 10.1073/pnas.2531786100
Membrane tube formation from giant vesicles by dynamic association of motor proteins
Abstract
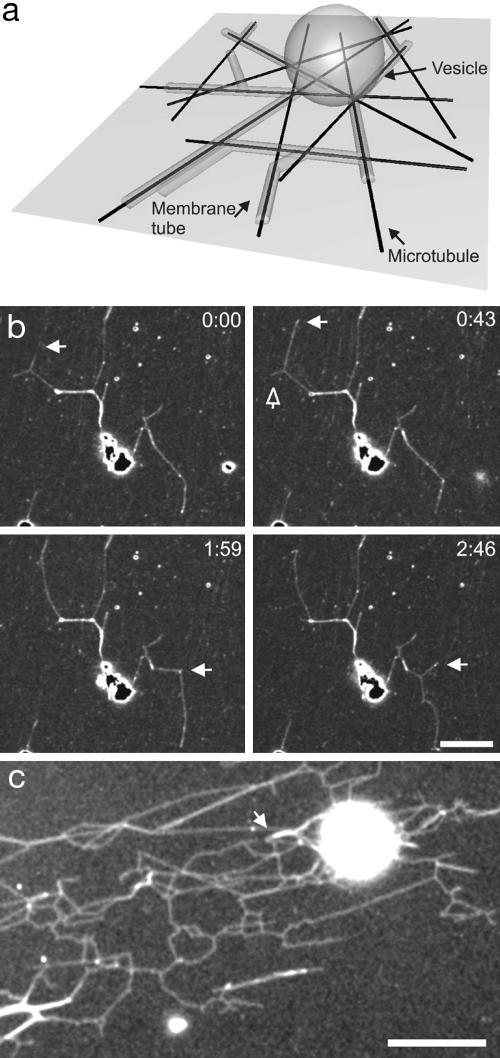
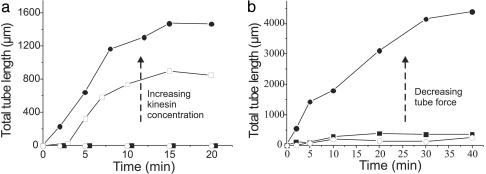
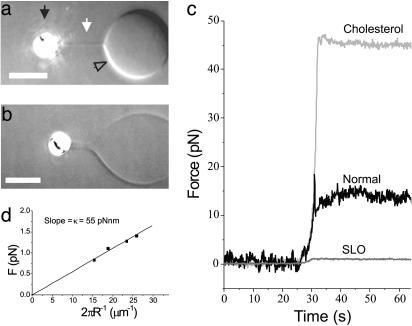
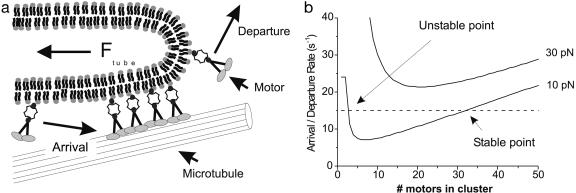
The tubular morphology of intracellular membranous compartments is actively maintained through interactions with motor proteins and the cytoskeleton. Moving along cytoskeletal elements, motor proteins exert forces on the membranes to which they are attached, resulting in the formation of membrane tubes and tubular networks. To study the formation of membrane tubes by motor proteins, we developed an in vitro assay consisting of purified kinesin proteins directly linked to the lipids of giant unilamellar vesicles. When the vesicles are brought into contact with a network of immobilized microtubules, membrane tubes and tubular networks are formed. Through systematic variation of the kinesin concentration and membrane composition we study the mechanism involved. We show that a threshold concentration of motor proteins is needed and that a low membrane tension facilitates tube formation. Forces involved in tube formation were measured directly with optical tweezers and are shown to depend only on the tension and bending rigidity of the membrane. The forces were found to be higher than can be generated by individual motor proteins, indicating that multiple motors were working together to pull tubes. We propose a simple mechanism by which individual motor proteins can dynamically associate into clusters that provide the force needed for the formation of tubes, explaining why, in contrast to earlier findings [Roux, A., Cappello, G., Cartaud, J., Prost, J., Goud, B. & Bassereau, P. (2002) Proc. Natl. Acad. Sci. USA 99, 5394-5399], motor proteins do not need to be physically linked to each other to be able to pull tubes.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
