

Abnormal surface liquid pH regulation by cultured cystic fibrosis bronchial epithelium
- PMID: 14668433
- PMCID: PMC307696
- DOI: 10.1073/pnas.2634339100
Abnormal surface liquid pH regulation by cultured cystic fibrosis bronchial epithelium
Abstract
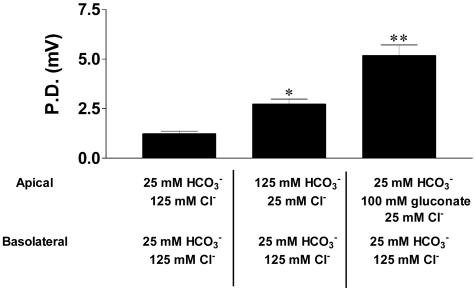
Cystic fibrosis (CF) transmembrane conductance regulator (CFTR)-dependent airway epithelial bicarbonate transport is hypothesized to participate in airway surface liquid pH regulation and contribute to lung defense. We measured pH and ionic composition in apical surface liquid (ASL) on polarized normal (NL) and CF primary bronchial epithelial cell cultures under basal conditions, after cAMP stimulation, and after challenge with luminal acid loads. Under basal conditions, CF epithelia acidified ASL more rapidly than NL epithelia. Two ASL pH regulatory paths that contributed to basal pH were identified in the apical membrane of airway epithelia, and their activities were measured. We detected a ouabain-sensitive (nongastric) H+,K+-ATPase that acidified ASL, but its activity was not different in NL and CF cultures. We also detected the following evidence for a CFTR-dependent HCO3- secretory pathway that was defective in CF: (i). ASL [HCO3-] was higher in NL than CF ASL; (ii). activating CFTR with forskolin/3-isobutyl-1-methylxanthine alkalinized NL ASL but acidified CF ASL; and (iii). NL airway epithelia more rapidly and effectively alkalinized ASL in response to a luminal acid challenge than CF epithelia. We conclude that cultured human CF bronchial epithelial pHASL is abnormally regulated under basal conditions because of absent CFTR-dependent HCO3- secretion and that this defect can lead to an impaired capacity to respond to airway conditions associated with acidification of ASL.
Figures





References
-
- Cheng, S. H., Gregory, R. J., Marshall, J., Paul, S., Souza, D. W., White, G. A., O'Riordan, C. R. & Smith, A. E. (1990) Cell 63, 827-834. - PubMed
-
- Kartner, N., Hanrahan, J. W., Jensen, T. J., Naismith, A. L., Sun, S. Z., Ackerley, C. A., Reyes, E. F., Tsui, L. C., Rommens, J. M., Bear, C. E., et al. (1991) Cell 64, 681-691. - PubMed
-
- Johansen, P. G., Anderson, C. M. & Hadorn, B. (1968) Lancet 1, 455-460. - PubMed
-
- Kopelman, H., Durie, P., Gaskin, K., Weizman, Z. & Forstner, G. (1985) N. Engl. J. Med. 312, 329-334. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
