

A striking quality control subcompartment in Saccharomyces cerevisiae: the endoplasmic reticulum-associated compartment
- PMID: 14668485
- PMCID: PMC329403
- DOI: 10.1091/mbc.e03-07-0546
A striking quality control subcompartment in Saccharomyces cerevisiae: the endoplasmic reticulum-associated compartment
Abstract
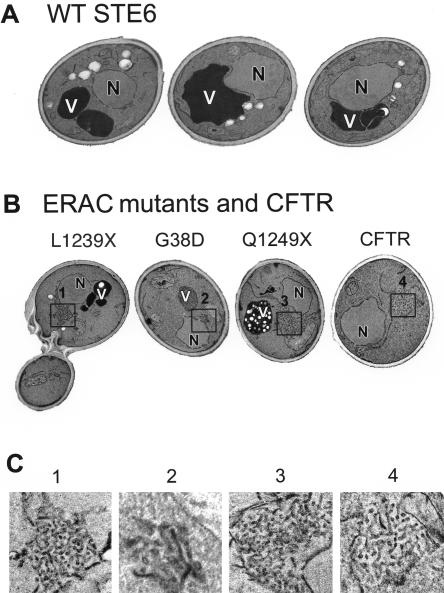
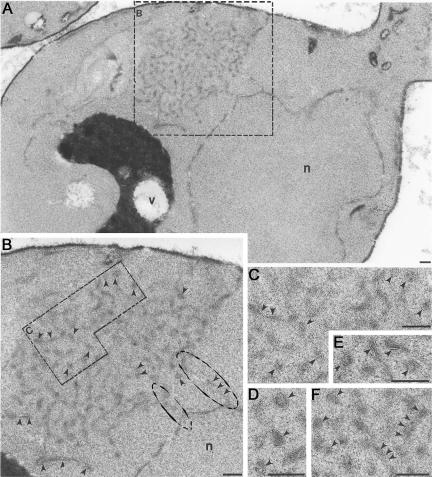
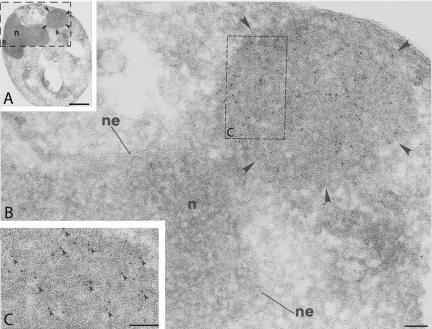
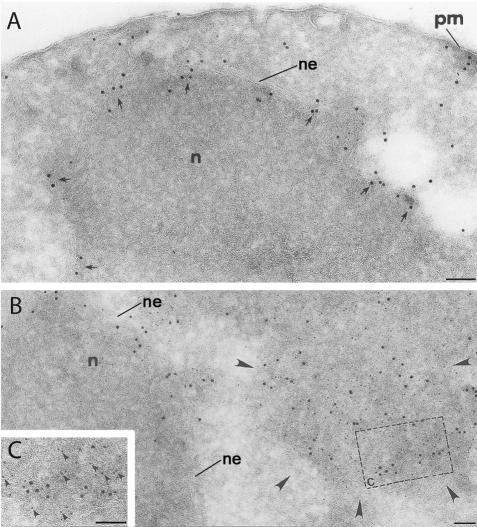
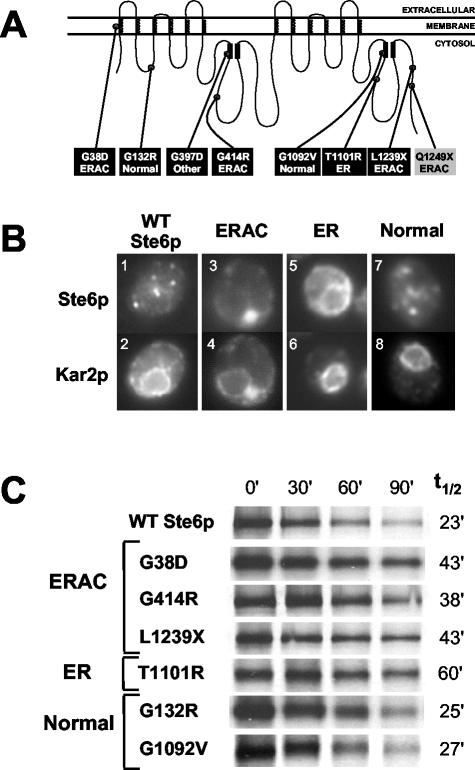
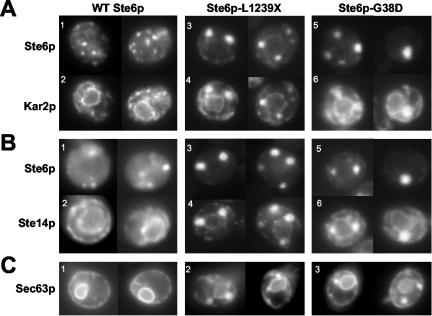
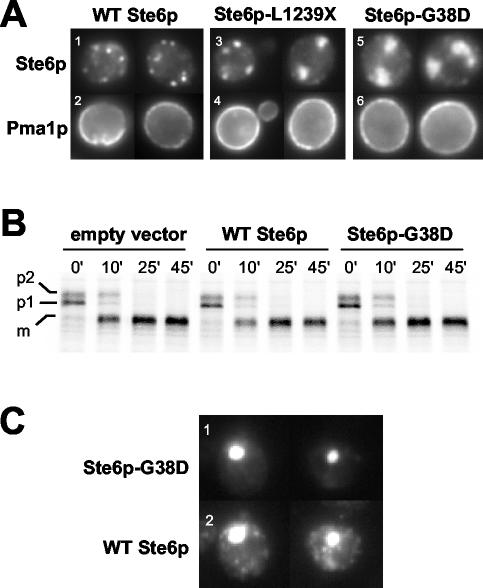
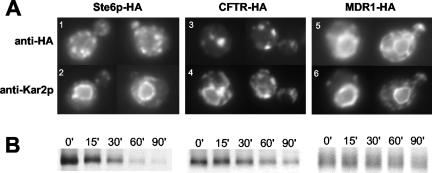
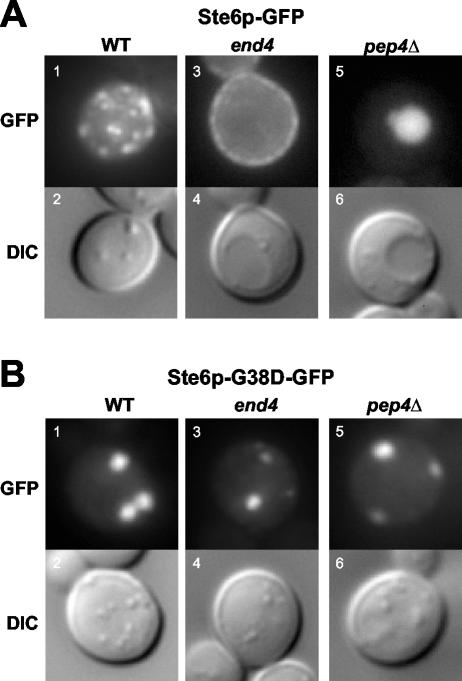
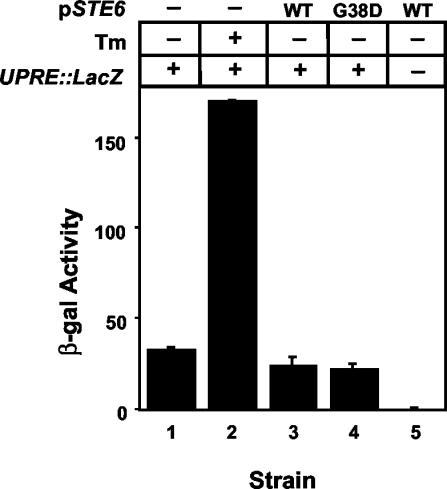
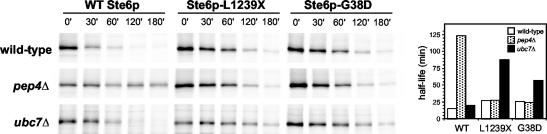
The folding of nascent secretory and membrane proteins is monitored by the endoplasmic reticulum (ER) quality control system. Misfolded proteins are retained in the ER and can be removed by ER-associated degradation. As a model for the ER quality control of multispanning membrane proteins in yeast, we have been studying mutant forms of Ste6p. Here, we identify mislocalized mutant forms of Ste6p that induce the formation of, and localize to, prominent structures that are absent in normal cells. We have named these structures ER-associated compartments (ERACs), based on their juxtaposition to and connection with the ER, as observed by fluorescence and electron microscopy. ERACs comprise a network of tubulo-vesicular structures that seem to represent proliferated ER membranes. Resident ER lumenal and membrane proteins are present in ERACs in addition to their normal ER localization, suggesting there is no barrier for their entry into ERACs. However, the forms of Ste6p in ERACs are excluded from the ER and do not enter the secretory pathway; instead, they are ultimately targeted for ER-associated degradation. The presence of ERACs does not adversely affect secretory protein traffic through the ER and does not lead to induction of the unfolded protein response. We propose that ERACs may be holding sites to which misfolded membrane proteins are specifically diverted so as not to interfere with normal cellular functions. We discuss the likelihood that related ER membrane proliferations that form in response to certain other mutant or unassembled membrane proteins may be substantially similar to ERACs.
Figures
References
-
- Ambudkar, S.V., Dey, S., Hrycyna, C.A., Ramachandra, M., Pastan, I., and Gottesman, M.M. (1999). Biochemical, cellular, and pharmacological aspects of the multidrug transporter. Annu. Rev. Pharmacol. Toxicol. 39, 361-398. - PubMed
-
- Barlowe, C. (2003). Signals for COPII-dependent export from the ER: what's the ticket out? Trends Cell Biol. 13, 295-300. - PubMed
-
- Berkower, C., Taglicht, D., and Michaelis, S. (1996). Functional and physical interactions between partial molecules of STE6, a yeast ATP-binding cassette protein. J. Biol. Chem. 271, 22983-22989. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
