

N-terminal domain of the murine coronavirus receptor CEACAM1 is responsible for fusogenic activation and conformational changes of the spike protein
- PMID: 14671103
- PMCID: PMC303413
- DOI: 10.1128/jvi.78.1.216-223.2004
N-terminal domain of the murine coronavirus receptor CEACAM1 is responsible for fusogenic activation and conformational changes of the spike protein
Abstract
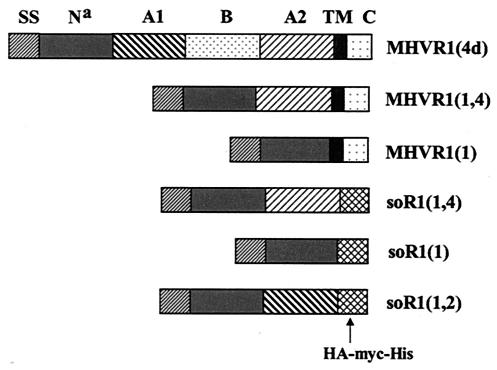
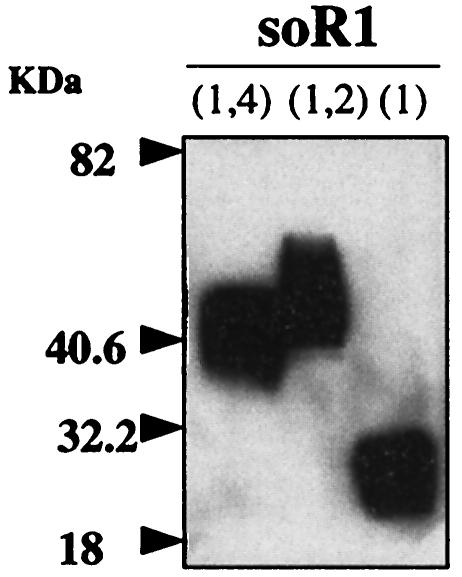
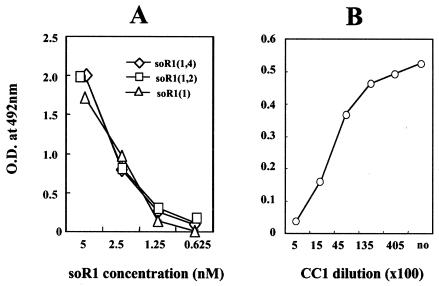
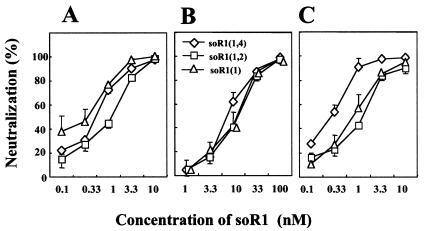
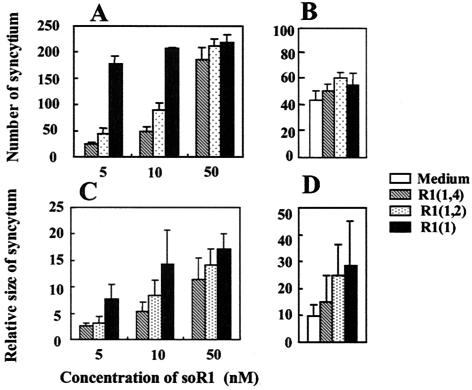
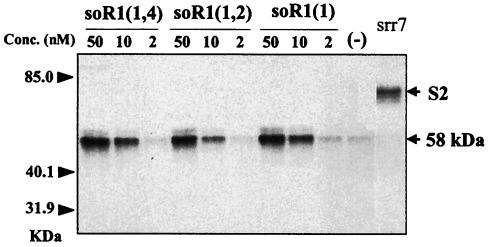
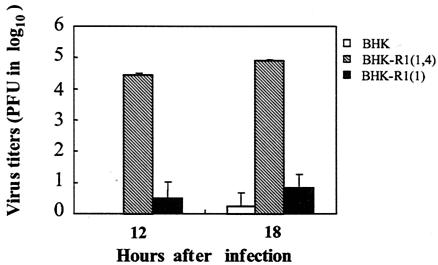
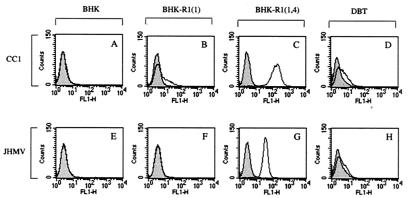
The mouse hepatitis virus (MHV) receptor (MHVR), CEACAM1, has two different functions for MHV entry into cells: binding to MHV spike protein (S protein) and activation of the S protein to execute virus-cell membrane fusion, the latter of which is accompanied by conformational changes of the S protein. The MHVR comprising the N-terminal and fourth domains [R1(1,4)] displays these two activities, and the N domain is thought to be critical for binding to MHV. In this study, we have addressed whether or not the N domain alone is sufficient for these activities. We examined three types of soluble form MHVR (soMHVR), one consisting of the N domain alone [soR1(1)], one with the N and second domains [soR1(1,2)], and one [soR1(1,4)] expressed by recombinant baculoviruses. We assessed the abilities of these three types of soMHVR to bind to MHV, activate fusogenicity, and induce conformational changes of the S protein. All three types of soMHVR similarly bound to MHV, as examined by a solid-phase binding assay and neutralized MHV infectivity. They also activated S protein fusogenicity and induced its conformational changes with similar levels of efficiency. However, R1(1) expressed on the BHK cell surface failed to serve as a receptor in spite of a sufficient level of expression. The inability of expressed R1(1) to work as a receptor was due to the inaccessibility of virions to R1(1); however, these were accessible using the MHVR-specific monoclonal antibody CC1. These results collectively indicated that the N domain retains all biological activities necessary for receptor function.
Figures
References
-
- Beauchemin, N., et al. 1999. Redefined nomenclature for members of the carcinoembryonic antigen family. Exp. Cell Res. 252:243-249. - PubMed
-
- Chen, D. C., and P. S. Kim. 1998. HIV entry and its inhibition. Cell 93:681-684. - PubMed
-
- Dalgleish, A. G., P. C. Beverley, P. R. Clapham, D. H. Crawford, M. F. Greaves, and R. A. Weiss. 1984. The CD4 (T4) antigen is an essential component of the receptor for the AIDS retrovirus. Nature 312:763-766. - PubMed
-
- Drosten, C., et al. 2003. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 348:1967-1976. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
