

doi: 10.1128/jvi.78.1.250-256.2004.
A JC virus-induced signal is required for infection of glial cells by a clathrin- and eps15-dependent pathway
Affiliations
- PMID: 14671106
- PMCID: PMC303400
- DOI: 10.1128/jvi.78.1.250-256.2004
Item in Clipboard
A JC virus-induced signal is required for infection of glial cells by a clathrin- and eps15-dependent pathway
J Virol.
2004 Jan.
Erratum in
-
Correction for Querbes et al., "A JC Virus-Induced Signal Is Required for Infection of Glial Cells by a Clathrin- and eps15-Dependent Pathway".J Virol. 2024 Apr 16;98(4):e0028224. doi: 10.1128/jvi.00282-24. Epub 2024 Mar 26. J Virol. 2024. PMID: 38529953 Free PMC article. No abstract available.
Abstract
Infectious entry of JC virus (JCV) into human glial cells occurs by receptor-mediated clathrin-dependent endocytosis. In this report we demonstrate that the tyrosine kinase inhibitor genistein blocks virus entry and inhibits infection. Transient expression of dominant-negative eps15 mutants, including a phosphorylation-defective mutant, inhibited both virus entry and infection. We also show that the JCV-induced signal activates the mitogen-activated protein kinases ERK1 and ERK2. These data demonstrate that JC virus binding to human glial cells induces an intracellular signal that is critical for entry and infection by a ligand-inducible clathrin-dependent mechanism.
Figures
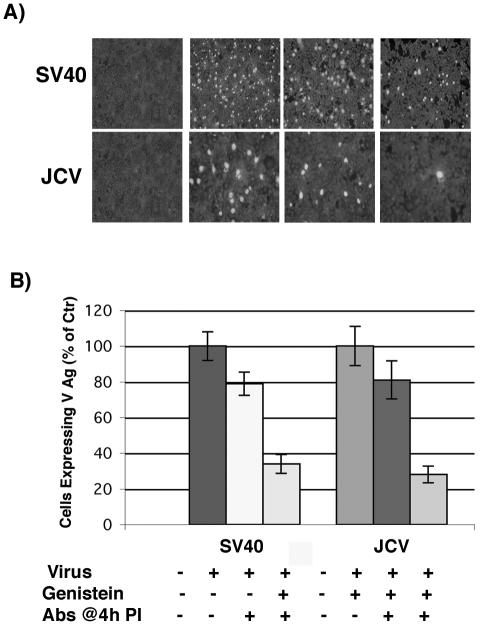
Genistein inhibits JCV and SV40 infection of glial cells. (A) SVG-A cells grown on coverslips were pretreated with 100 μM genistein for 1 h at 37°C. Cells were then infected with JCV or SV40 in the continued presence of genistein for 4 h at 37°C, and anti-JCV or anti-SV40 neutralizing antibodies were added at 4 h postinfection to neutralize any remaining extracellular virus. At 72 h postinfection the cells were washed and fixed, and infection was scored by an indirect immunofluorescence assay for the late viral protein VP1. (B) VP1-positive cells were scored by counting, and the results are graphed.
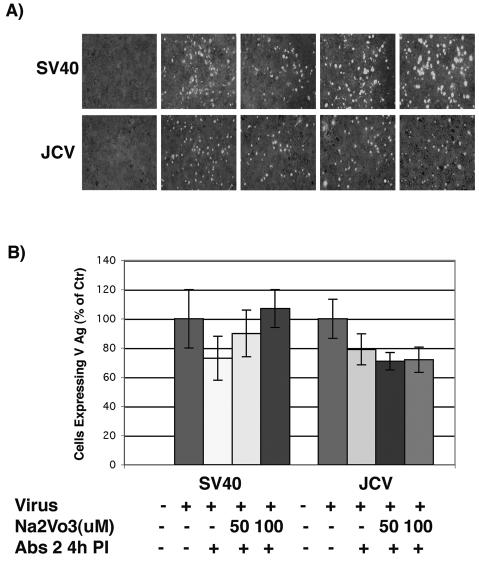
Sodium orthovanadate does not inhibit JCV infection of glial cells. (A) SVG-A cells grown on coverslips were pretreated with 50 or 100 μM sodium orthovanadate for 1 h at 37°C. Cells were then infected with JCV or SV40 in the continued presence of NaVO3 for 4 h at 37°C, and anti-JCV or anti-SV40 neutralizing antibodies were added at 4 h postinfection to neutralize any remaining extracellular virus. At 72 h postinfection the cells were washed and fixed, and infection was scored by an indirect immunofluorescence assay for the late viral protein VP1. (B) VP1-positive cells were scored by counting, and the results are graphed.
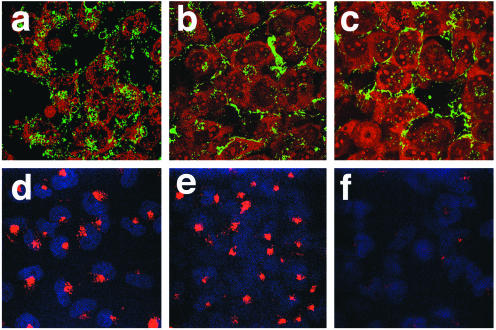
Genistein and chlorpromazine inhibit entry of FITC-labeled JCV. SVG-A cells grown on coverslips were either untreated (control) (a and d) or pretreated with 100 μM genistein (b and e) or 25 μM chlorpromazine (c and f) for 1 h. The cells were then incubated with FITC-labeled JCV (a to c) for 4 h in the continued presence of drug or incubated with 35 μg of rhodamine-labeled transferrin/ml (d to f) for 30 min also in the continued presence of the drug. Cells were washed and fixed in 4% paraformaldehyde, and coverslips were mounted on slides with mounting medium containing propidium iodide (a to c) or DAPI (d to f). Slides were viewed using a laser scanning confocal microscope with a 63× objective.
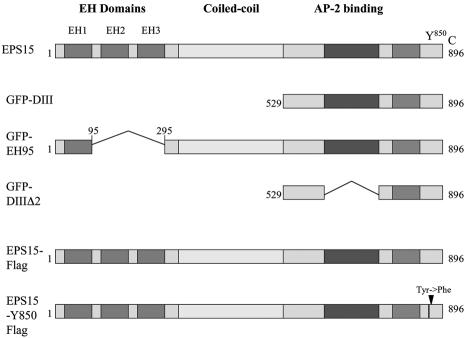
Schematic representation of full-length eps15 and the dominant-negative mutant constructs used in these experiments. eps15 has an amino terminus containing three protein-protein interaction EH domains, a central coiled-coil domain thought to be involved in homodimerization, and a carboxyl terminus involved in association with the clathrin adaptor protein AP-2 during clathrin pit formation. The DIII (D3) and EH95 constructs are dominant-negative inhibitors of eps15 function and inhibit clathrin-dependent endocytosis, whereas the DIIIΔ2 (D3D2) construct serves as a negative control and has no effect on clathrin-dependent endocytosis. The FLAG constructs are either a control full-length eps15 construct or full-length eps15 with a point mutation (Tyr→Phe) in the major tyrosine phosphorylation site, Y850, which has been shown to block EGF internalization but not transferrin internalization.
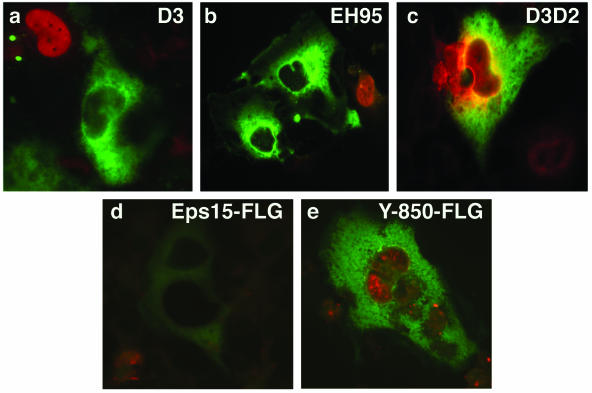
Dominant-negative eps15 constructs inhibit JCV infection. SVG-A cells grown on coverslips were transfected with the indicated GFP- or FLAG-tagged dominant-negative eps15 constructs or control constructs for 24 h. Cells were then infected with JCV, and at 72 h postinfection the cells were fixed and stained with an anti-V antigen (VP1) monoclonal antibody (red). The GFP-expressing constructs were directly visualized, and the FLAG-tagged constructs were visualized using anti-FLAG antibody and an Alexa-Fluor 488-labeled secondary antibody. JCV was unable to infect any of the cells expressing a dominant-negative construct (EH95, D3, and eps15-Y850F). In contrast, JCV readily infected cells expressing control constructs (D3D2 and eps15wt).
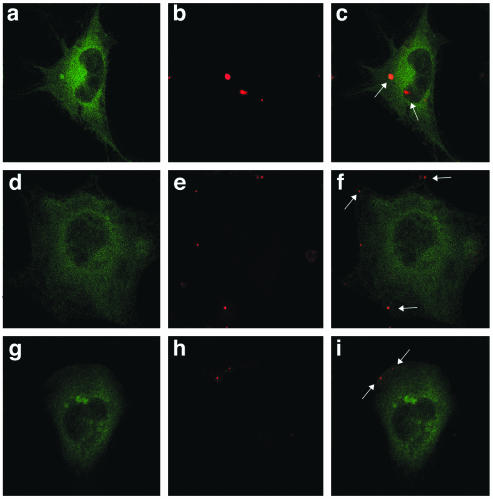
Dominant-negative eps15 constructs block JCV entry. SVG-A cells grown on coverslips were transfected with either D3 (a to c) or EH95 (d and e) dominant-negative eps15-GFP constructs or the D3Δ2 control construct (f to h) for 24 h. Twenty-four hours posttransfection, cells were exposed to Alexa-Fluor 594-labeled JCV (red) for 4 h at 37°C and then fixed in 4% paraformaldehyde for 30 min. The cells were mounted using fluorescent mounting medium and viewed using a confocal microscope. Arrows indicate the presence of internalized virus (c) or membrane-bound virus (f and i).
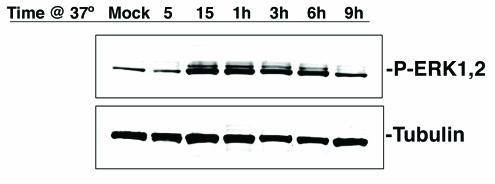
JCV activates the MAPKs ERK1 and ERK2. SVG-A cells serum starved overnight were incubated with JCV or just medium (mock) for 30 min on ice, and then cells were shifted to 37°C to allow virus internalization. Lysates were collected at the indicated time points following the shift to 37°C, and samples were run on 4- to-15% gradient gels, transferred to nitrocellulose, and blotted with antibodies recognizing the phosphorylated or active form of ERK1 and -2 or with antibodies to tubulin as a loading control.
Similar articles
-
JC virus enters human glial cells by clathrin-dependent receptor-mediated endocytosis.J Virol. 2000 Mar;74(5):2288-92. doi: 10.1128/jvi.74.5.2288-2292.2000. J Virol. 2000. PMID: 10666259 Free PMC article.
-
Ligand-induced clathrin-mediated endocytosis of the keratinocyte growth factor receptor occurs independently of either phosphorylation or recruitment of eps15.FEBS Lett. 2003 Oct 23;553(3):262-70. doi: 10.1016/s0014-5793(03)01020-2. FEBS Lett. 2003. PMID: 14572635
-
Mouse hepatitis virus type 2 enters cells through a clathrin-mediated endocytic pathway independent of Eps15.J Virol. 2008 Aug;82(16):8112-23. doi: 10.1128/JVI.00837-08. Epub 2008 Jun 11. J Virol. 2008. PMID: 18550663 Free PMC article.
-
Epidermal growth factor pathway substrate 15, Eps15.Int J Biochem Cell Biol. 1999 Aug;31(8):805-9. doi: 10.1016/s1357-2725(99)00042-4. Int J Biochem Cell Biol. 1999. PMID: 10481267 Review.
-
Molecular mechanisms underlying endocytosis and sorting of ErbB receptor tyrosine kinases.FEBS Lett. 2001 Feb 16;490(3):142-52. doi: 10.1016/s0014-5793(01)02117-2. FEBS Lett. 2001. PMID: 11223029 Review.
Cited by
-
Japanese encephalitis virus infects porcine kidney epithelial PK15 cells via clathrin- and cholesterol-dependent endocytosis.Virol J. 2013 Aug 12;10:258. doi: 10.1186/1743-422X-10-258. Virol J. 2013. PMID: 23937769 Free PMC article.
-
CRISPR antiviral inhibits neurotrophic JC polyomavirus in 2D and 3D culture models through dual-gRNA excision by SaCas9.Mol Ther Nucleic Acids. 2025 May 14;36(2):102556. doi: 10.1016/j.omtn.2025.102556. eCollection 2025 Jun 10. Mol Ther Nucleic Acids. 2025. PMID: 40510594 Free PMC article.
-
Adipocyte Plasma Membrane Protein (APMAP) promotes JC Virus (JCPyV) infection in human glial cells.Virology. 2020 Sep;548:17-24. doi: 10.1016/j.virol.2020.06.002. Epub 2020 Jun 7. Virology. 2020. PMID: 32838939 Free PMC article.
-
Rearrangement in the Hypervariable Region of JC Polyomavirus Genomes Isolated from Patient Samples and Impact on Transcription Factor-Binding Sites and Disease Outcomes.Int J Mol Sci. 2022 May 20;23(10):5699. doi: 10.3390/ijms23105699. Int J Mol Sci. 2022. PMID: 35628509 Free PMC article.
-
The oncogenic roles of JC polyomavirus in cancer.Front Oncol. 2022 Sep 23;12:976577. doi: 10.3389/fonc.2022.976577. eCollection 2022. Front Oncol. 2022. PMID: 36212474 Free PMC article. Review.
References
-
- Antinori, A., A. Ammassari, M. L. Giancola, A. Cingolani, S. Grisetti, R. Murri, L. Alba, B. Ciancio, F. Soldani, D. Larussa, G. Ippolito, and A. De Luca. 2001. Epidemiology and prognosis of AIDS-associated progressive multifocal leukoencephalopathy in the HAART era. J. Neurovirol. 7:323-328. - PubMed
-
- Benmerah, A., M. Bayrou, N. Cerf-Bensussan, and A. Dautry-Varsat. 1999. Inhibition of clathrin-coated pit assembly by an Eps15 mutant. J. Cell Sci. 112:1303-1311. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
