

Ultrafast folding of alpha3D: a de novo designed three-helix bundle protein
- PMID: 14671331
- PMCID: PMC307594
- DOI: 10.1073/pnas.2136623100
Ultrafast folding of alpha3D: a de novo designed three-helix bundle protein
Abstract
Here, we describe the folding/unfolding kinetics of alpha3D, a small designed three-helix bundle. Both IR temperature jump and ultrafast fluorescence mixing methods reveal a single-exponential process consistent with a minimal folding time of 3.2 +/- 1.2 micros (at approximately 50 degrees C), indicating that a protein can fold on the 1- to 5-micros time scale. Furthermore, the single-exponential nature of the relaxation indicates that the prefactor for transition state (TS)-folding models is probably >or=1 (micros)-1 for a protein of this size and topology. Molecular dynamics simulations and IR spectroscopy provide a molecular rationale for the rapid, single-exponential folding of this protein. alpha3D shows a significant bias toward local helical structure in the thermally denatured state. The molecular dynamics-simulated TS ensemble is highly heterogeneous and dynamic, allowing access to the TS via multiple pathways.
Figures

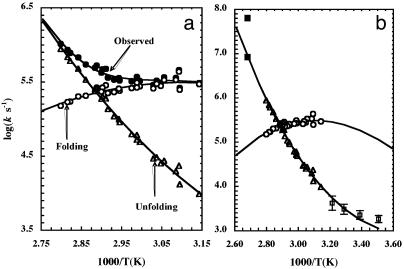
 Os. (b) Difference IR spectra generated by subtracting the spectrum collected at 2.5°C from the spectra in a. Arrows indicate the direction of changes when temperature is increased. (c) Relative band areas of the three amide bands as a function of temperature.
Os. (b) Difference IR spectra generated by subtracting the spectrum collected at 2.5°C from the spectra in a. Arrows indicate the direction of changes when temperature is increased. (c) Relative band areas of the three amide bands as a function of temperature.


Similar articles
-
Kinetics and motional dynamics of spin-labeled yeast iso-1-cytochrome c: 1. Stopped-flow electron paramagnetic resonance as a probe for protein folding/unfolding of the C-terminal helix spin-labeled at cysteine 102.Biochemistry. 1997 Mar 11;36(10):2884-97. doi: 10.1021/bi962155i. Biochemistry. 1997. PMID: 9062118
-
Kinetics and thermodynamics of folding of a de novo designed four-helix bundle protein.J Mol Biol. 1996 Oct 25;263(2):323-43. doi: 10.1006/jmbi.1996.0578. J Mol Biol. 1996. PMID: 8913310
-
Simulation and experiment at high temperatures: ultrafast folding of a thermophilic protein by nucleation-condensation.J Mol Biol. 2005 Apr 8;347(4):855-70. doi: 10.1016/j.jmb.2004.12.061. J Mol Biol. 2005. PMID: 15769475
-
Simulating the peptide folding kinetic related spectra based on the Markov State Model.Adv Exp Med Biol. 2014;805:199-220. doi: 10.1007/978-3-319-02970-2_9. Adv Exp Med Biol. 2014. PMID: 24446363 Review.
-
Molecular dynamics simulations of the protein unfolding/folding reaction.Acc Chem Res. 2002 Jun;35(6):422-9. doi: 10.1021/ar0100834. Acc Chem Res. 2002. PMID: 12069627 Review.
Cited by
-
Folding and Stabilization of Native-Sequence-Reversed Proteins.Sci Rep. 2016 Apr 26;6:25138. doi: 10.1038/srep25138. Sci Rep. 2016. PMID: 27113844 Free PMC article.
-
Full reconstruction of a vectorial protein folding pathway by atomic force microscopy and molecular dynamics simulations.J Biol Chem. 2010 Dec 3;285(49):38167-72. doi: 10.1074/jbc.M110.179697. Epub 2010 Sep 24. J Biol Chem. 2010. PMID: 20870713 Free PMC article.
-
Probing the folding intermediate of Rd-apocyt b562 by protein engineering and infrared T-jump.Protein Sci. 2007 Jun;16(6):1176-83. doi: 10.1110/ps.062505607. Epub 2007 May 1. Protein Sci. 2007. PMID: 17473017 Free PMC article.
-
Sculpting Metal-binding Environments in De Novo Designed Three-helix Bundles.Isr J Chem. 2015 Jan;55(1):85-95. doi: 10.1002/ijch.201400146. Epub 2015 Jan 15. Isr J Chem. 2015. PMID: 29353917 Free PMC article.
-
Microsecond folding experiments and simulations: a match is made.Phys Chem Chem Phys. 2013 Mar 14;15(10):3372-88. doi: 10.1039/c3cp43992e. Epub 2013 Jan 29. Phys Chem Chem Phys. 2013. PMID: 23361200 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- CA06927/CA/NCI NIH HHS/United States
- GM-54616/GM/NIGMS NIH HHS/United States
- R01 GM065978/GM/NIGMS NIH HHS/United States
- GM-065978/GM/NIGMS NIH HHS/United States
- GM-056250/GM/NIGMS NIH HHS/United States
- R01 GM056250/GM/NIGMS NIH HHS/United States
- R01 GM054616/GM/NIGMS NIH HHS/United States
- P41 RR001348/RR/NCRR NIH HHS/United States
- GM-058079/GM/NIGMS NIH HHS/United States
- R37 GM054616/GM/NIGMS NIH HHS/United States
- P30 CA006927/CA/NCI NIH HHS/United States
- 5P41-RR01348/RR/NCRR NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
