

12-lipoxygenase metabolites of arachidonic acid mediate metabotropic glutamate receptor-dependent long-term depression at hippocampal CA3-CA1 synapses
- PMID: 14673007
- PMCID: PMC6740529
- DOI: 10.1523/JNEUROSCI.23-36-11427.2003
12-lipoxygenase metabolites of arachidonic acid mediate metabotropic glutamate receptor-dependent long-term depression at hippocampal CA3-CA1 synapses
Abstract
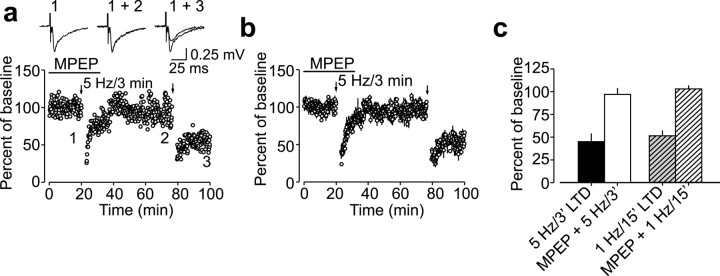
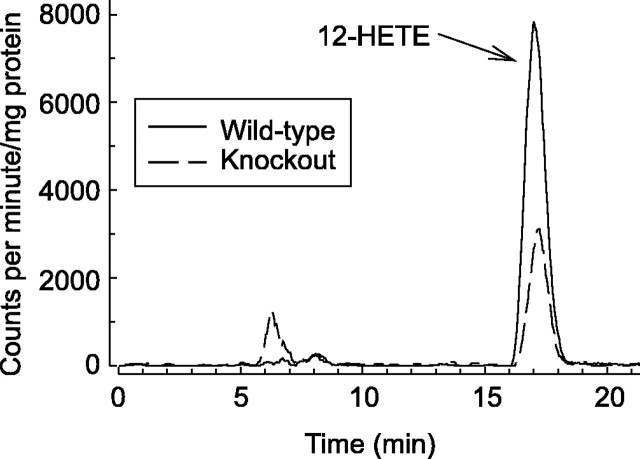
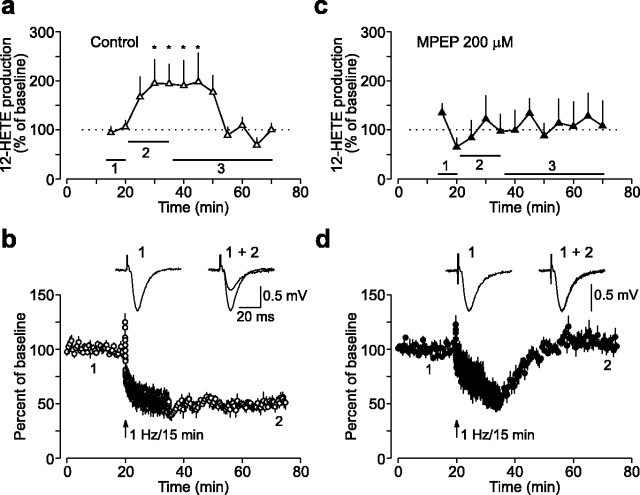
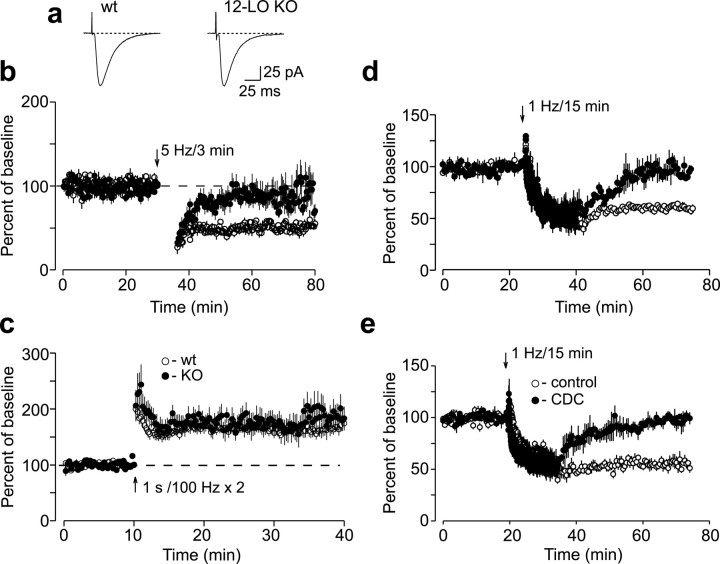
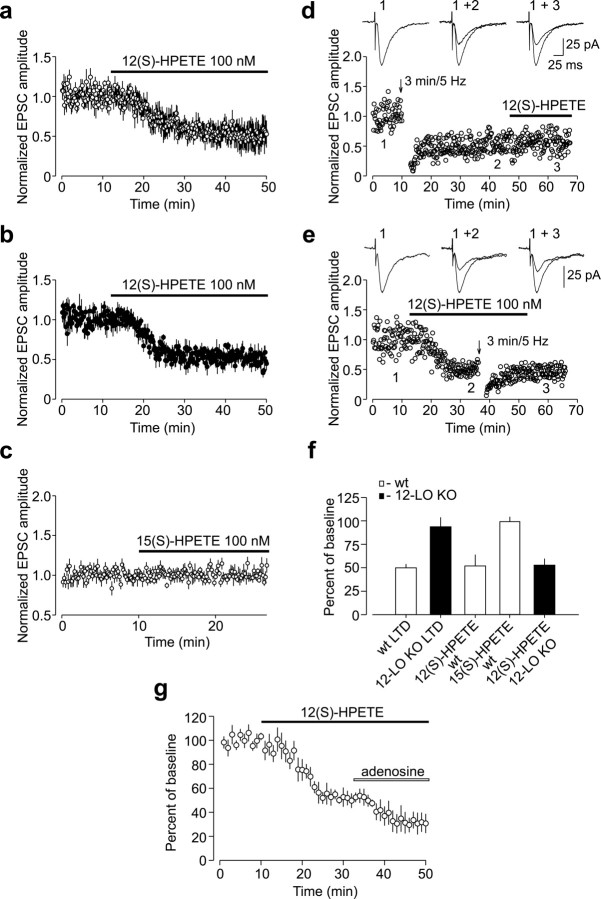
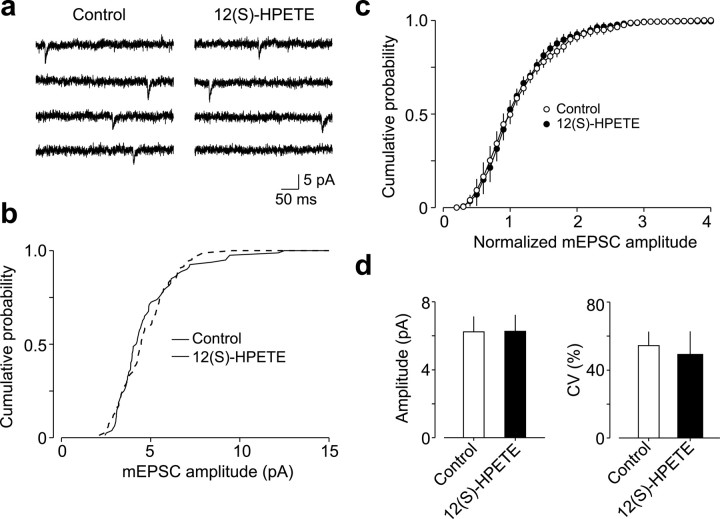
Arachidonic acid metabolites have been proposed as signaling molecules in hippocampal long-term potentiation (LTP) and long-term depression (LTD) for >15 years. However, the functional role of these molecules remains controversial. Here we used a multidisciplinary biochemical, electrophysiological, and genetic approach to examine the function of the 12-lipoxygenase metabolites of arachidonic acid in long-term synaptic plasticity at CA3-CA1 synapses. We found that the 12-lipoxygenase pathway is required for the induction of metabotropic glutamate receptor-dependent LTD (mGluR-LTD), but is not required for LTP: (1) Hippocampal homogenates were capable of synthesizing the 12-lipoxygenase metabolite of arachidonic acid, 12(S)-hydroxyeicosa-5Z,8Z,10E,14Z-tetraenoic acid (HETE). (2) Stimulation protocols that induce mGluR-LTD lead to a release of 12-(S)-HETE from acute hippocampal slices. (3) A mouse in which the leukocyte-type 12-lipoxygenase (the neuronal isoform) was deleted through homologous recombination was deficient in mGluR-LTD, but showed normal LTP. (4) Pharmacological inhibition of 12-lipoxygenase also blocked induction of mGluR-LTD. (5) Finally, direct application of 12(S)-HPETE, but not 15(S)-HPETE, to hippocampal slices induced a long-term depression of synaptic transmission that mimicked and occluded mGluR-LTD induced by synaptic stimulation. Thus, 12(S)-hydroperoxyeicosa-5Z, 8Z, 10E, 14Z-tetraenoic acid (12(S)-HPETE), a 12-lipoxygenase metabolite of arachidonic acid, satisfies all of the criteria of a messenger molecule that is actively recruited for the induction of mGluR-LTD.
Figures
Similar articles
-
Association of mGluR-Dependent LTD of Excitatory Synapses with Endocannabinoid-Dependent LTD of Inhibitory Synapses Leads to EPSP to Spike Potentiation in CA1 Pyramidal Neurons.J Neurosci. 2019 Jan 9;39(2):224-237. doi: 10.1523/JNEUROSCI.2935-17.2018. Epub 2018 Nov 20. J Neurosci. 2019. PMID: 30459224 Free PMC article.
-
GABA(A) receptor inhibition does not affect mGluR-dependent LTD at hippocampal Schaffer collateral-CA1 synapses.Neurosci Lett. 2009 Dec 18;467(1):20-5. doi: 10.1016/j.neulet.2009.09.053. Epub 2009 Oct 1. Neurosci Lett. 2009. PMID: 19800390
-
12-Lipoxygenase regulates hippocampal long-term potentiation by modulating L-type Ca2+ channels.J Neurosci. 2010 Feb 3;30(5):1822-31. doi: 10.1523/JNEUROSCI.2168-09.2010. J Neurosci. 2010. PMID: 20130191 Free PMC article.
-
Metabotropic glutamate receptor-mediated long-term depression: molecular mechanisms.Pharmacol Rev. 2009 Dec;61(4):395-412. doi: 10.1124/pr.109.001735. Epub 2009 Nov 19. Pharmacol Rev. 2009. PMID: 19926678 Free PMC article. Review.
-
Do group I metabotropic glutamate receptors mediate LTD?Neurobiol Learn Mem. 2017 Feb;138:85-97. doi: 10.1016/j.nlm.2016.08.010. Epub 2016 Aug 18. Neurobiol Learn Mem. 2017. PMID: 27545442 Review.
Cited by
-
Antipurinergic therapy corrects the autism-like features in the Fragile X (Fmr1 knockout) mouse model.Mol Autism. 2015 Jan 13;6:1. doi: 10.1186/2040-2392-6-1. eCollection 2015. Mol Autism. 2015. PMID: 25705365 Free PMC article.
-
Tyrosine phosphatases regulate AMPA receptor trafficking during metabotropic glutamate receptor-mediated long-term depression.J Neurosci. 2006 Mar 1;26(9):2544-54. doi: 10.1523/JNEUROSCI.4322-05.2006. J Neurosci. 2006. PMID: 16510732 Free PMC article.
-
Transcriptional regulation of β-secretase-1 by 12/15-lipoxygenase results in enhanced amyloidogenesis and cognitive impairments.Ann Neurol. 2012 Jan;71(1):57-67. doi: 10.1002/ana.22625. Ann Neurol. 2012. Retraction in: Ann Neurol. 2025 Mar;97(3):607. doi: 10.1002/ana.27184. PMID: 22275252 Free PMC article. Retracted.
-
Long-term potentiation in hippocampal oriens interneurons: postsynaptic induction, presynaptic expression and evaluation of candidate retrograde factors.Philos Trans R Soc Lond B Biol Sci. 2013 Dec 2;369(1633):20130133. doi: 10.1098/rstb.2013.0133. Print 2014 Jan 5. Philos Trans R Soc Lond B Biol Sci. 2013. PMID: 24298136 Free PMC article.
-
Developmental regulation of protein interacting with C kinase 1 (PICK1) function in hippocampal synaptic plasticity and learning.Proc Natl Acad Sci U S A. 2010 Dec 14;107(50):21784-9. doi: 10.1073/pnas.1016103107. Epub 2010 Nov 24. Proc Natl Acad Sci U S A. 2010. PMID: 21106762 Free PMC article.
References
-
- Bolshakov VY, Siegelbaum SA ( 1994) Postsynaptic induction and presynaptic expression of hippocampal long-term depression. Science 264: 1148-1152. - PubMed
-
- Bolshakov VY, Siegelbaum SA ( 1995) Hippocampal long-term depression: arachidonic acid as a potential retrograde messenger. Neuropharmacology 34: 1581-1587. - PubMed
-
- Bolshakov VY, Carboni L, Cobb M, Siegelbaum SA, Belardetti F ( 2000) Dual MAP kinase pathways mediate opposing forms of long-term plasticity at CA3-CA1 synapses. Nat Neurosci 3: 1107-1113. - PubMed
-
- Borsch-Haubold AG, Kramer RM, Watson SP ( 1997) Phosphorylation and activation of cytosolic phospholipase A2 by 38-kDa mitogen-activated protein kinase in collagen-stimulated human platelets. Eur J Biochem 245: 751-759. - PubMed
-
- Buttner N, Siegelbaum SA, Volterra A ( 1989) Direct modulation of Aplysia S-K+ channels by a 12-lipoxygenase metabolite of arachidonic acid. Nature 342: 553-555. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous