

CoAA, a nuclear receptor coactivator protein at the interface of transcriptional coactivation and RNA splicing
- PMID: 14673176
- PMCID: PMC303353
- DOI: 10.1128/MCB.24.1.442-453.2004
CoAA, a nuclear receptor coactivator protein at the interface of transcriptional coactivation and RNA splicing
Abstract
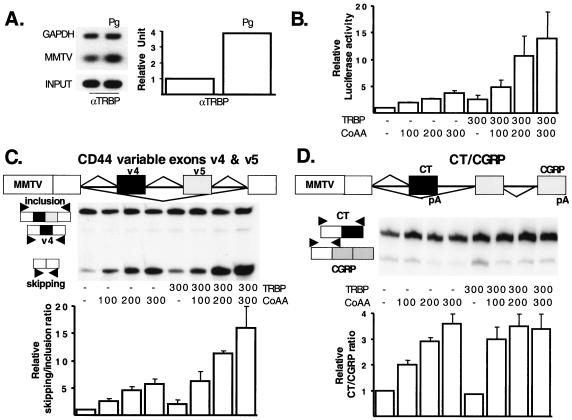
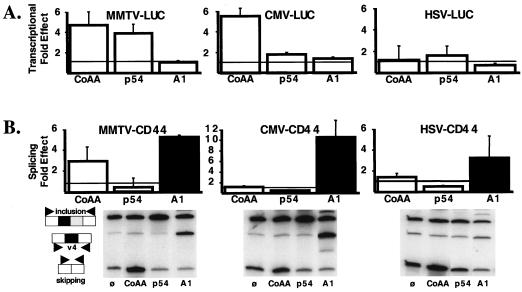
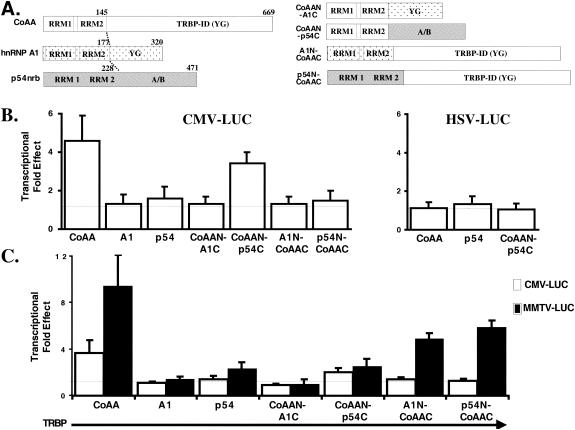
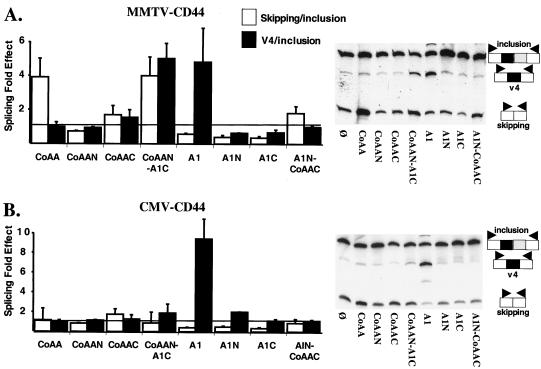
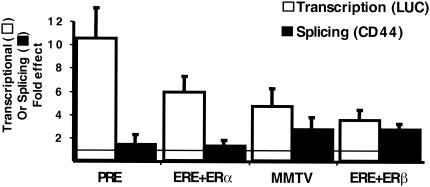
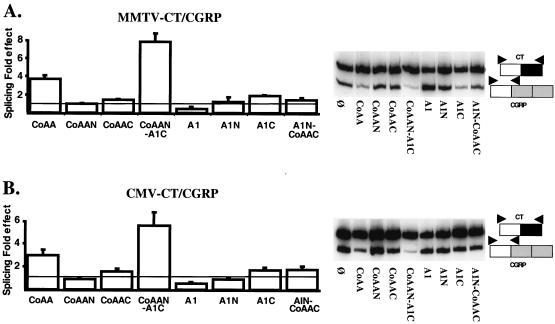
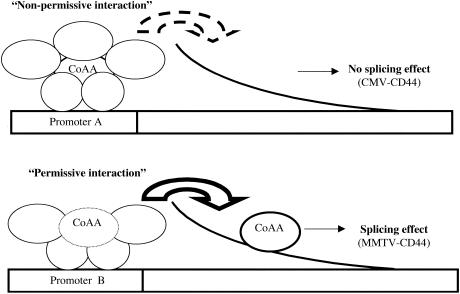
We have shown that steroid hormones coordinately control gene transcriptional activity and splicing decisions in a promoter-dependent manner. Our hypothesis is that a subset of hormonally recruited coregulators involved in regulation of promoter transcriptional activity also directly participate in alternative RNA splicing decisions. To gain insight into the molecular mechanisms by which transcriptional coregulators could control splicing decisions, we focused our attention on a recently identified coactivator, CoAA. This heterogeneous nuclear ribonucleoprotein (hnRNP)-like protein interacts with the transcriptional coregulator TRBP, a protein recruited to target promoters through interactions with activated nuclear receptors. Using transcriptional and splicing reporter genes driven by different promoters, we observed that CoAA mediates transcriptional and splicing effects in a promoter-preferential manner. We compared the activity of CoAA to the activity of other hnRNP-related proteins that, like CoAA, contain two N-terminal RNA recognition motifs (RRMs) followed by a C-terminal auxiliary domain and either have or have not been implicated in transcriptional control. By swapping either CoAA RRMs or the CoAA auxiliary domain with the corresponding domains of the proteins selected, we showed that depending on the promoter, the RRMs and the auxiliary domain of CoAA are differentially engaged in transcription. This contributes to the promoter-preferential effects mediated by CoAA on RNA splicing during the course of steroid hormone action.
Figures
References
-
- Aranda, A., and A. Pascual. 2001. Nuclear hormone receptors and gene expression. Physiol Rev. 81:1269-1304. - PubMed
-
- Auboeuf, D., A. Honig, S. M. Berget, and B. W. O'Malley. 2002. Coordinate regulation of transcription and splicing by steroid receptor coregulators. Science 298:416-419. - PubMed
-
- Caira, F., P. Antonson, M. Pelto-Huikko, E. Treuter, and J. A. Gustafsson. 2000. Cloning and characterization of RAP250, a novel nuclear receptor coactivator. J. Biol. Chem. 275:5308-5317. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases