

Free-energy landscapes of ion-channel gating are malleable: changes in the number of bound ligands are accompanied by changes in the location of the transition state in acetylcholine-receptor channels
- PMID: 14674774
- PMCID: PMC1463891
- DOI: 10.1021/bi0354334
Free-energy landscapes of ion-channel gating are malleable: changes in the number of bound ligands are accompanied by changes in the location of the transition state in acetylcholine-receptor channels
Abstract
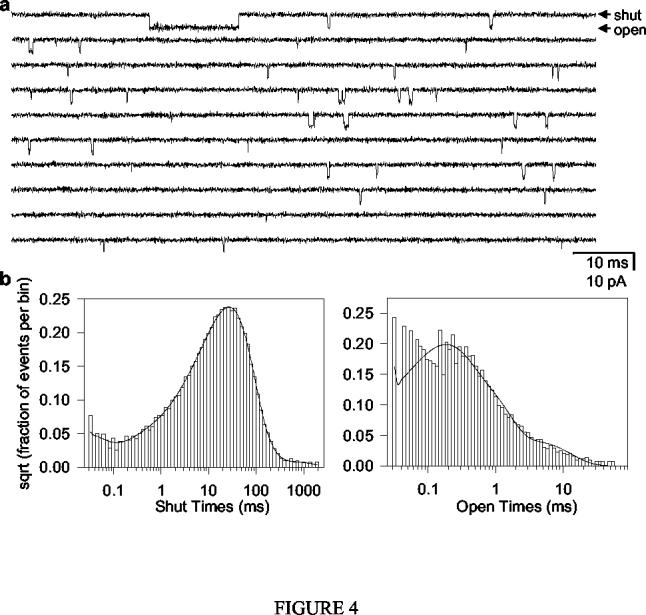
Acetylcholine-receptor channels (AChRs) are allosteric membrane proteins that mediate synaptic transmission by alternatively opening and closing ("gating") a cation-selective transmembrane pore. Although ligand binding is not required for the channel to open, the binding of agonists (for example, acetylcholine) increases the closed right harpoon over left harpoon open equilibrium constant because the ion-impermeable --> ion-permeable transition of the ion pathway is accompanied by a low-affinity --> high-affinity change at the agonist-binding sites. The fact that the gating conformational change of muscle AChRs can be kinetically modeled as a two-state reaction has paved the way to the experimental characterization of the corresponding transition state, which represents a snapshot of the continuous sequence of molecular events separating the closed and open states. Previous studies of fully (di) liganded AChRs, combining single-channel kinetic measurements, site-directed mutagenesis, and data analysis in the framework of the linear free-energy relationships of physical organic chemistry, have suggested a transition-state structure that is consistent with channel opening being an asynchronous conformational change that starts at the extracellular agonist-binding sites and propagates toward the intracellular end of the pore. In this paper, I characterize the gating transition state of unliganded AChRs, and report a remarkable difference: unlike that of diliganded gating, the unliganded transition state is not a hybrid of the closed- and open-state structures but, rather, is almost indistinguishable from the open state itself. This displacement of the transition state along the reaction coordinate obscures the mechanism underlying the unliganded closed right harpoon over left harpoon open reaction but brings to light the malleable nature of free-energy landscapes of ion-channel gating.
Figures







Similar articles
-
Structure of the transition state of gating in the acetylcholine receptor channel pore: a phi-value analysis.Biochemistry. 2002 Apr 30;41(17):5548-55. doi: 10.1021/bi011864f. Biochemistry. 2002. PMID: 11969415 Free PMC article.
-
Dynamics of the acetylcholine receptor pore at the gating transition state.Proc Natl Acad Sci U S A. 2005 Oct 18;102(42):15069-74. doi: 10.1073/pnas.0505090102. Epub 2005 Oct 10. Proc Natl Acad Sci U S A. 2005. PMID: 16217024 Free PMC article.
-
The role of loop 5 in acetylcholine receptor channel gating.J Gen Physiol. 2003 Nov;122(5):521-39. doi: 10.1085/jgp.200308885. Epub 2003 Oct 13. J Gen Physiol. 2003. PMID: 14557402 Free PMC article.
-
Acetylcholine receptors, between closed and open.Novartis Found Symp. 2002;245:223-34; discussion 234-9, 261-4. Novartis Found Symp. 2002. PMID: 12027011 Review.
-
Life at the top: the transition state of AChR gating.Sci STKE. 2003 Jun 24;2003(188):re11. doi: 10.1126/stke.2003.188.re11. Sci STKE. 2003. PMID: 12824477 Review.
Cited by
-
Energetics of gating at the apo-acetylcholine receptor transmitter binding site.J Gen Physiol. 2010 Apr;135(4):321-31. doi: 10.1085/jgp.200910384. J Gen Physiol. 2010. PMID: 20351060 Free PMC article.
-
Estimating binding affinities of the nicotinic receptor for low-efficacy ligands using mixtures of agonists and two-dimensional concentration-response relationships.J Gen Physiol. 2006 Jun;127(6):719-35. doi: 10.1085/jgp.200509438. J Gen Physiol. 2006. PMID: 16735756 Free PMC article.
-
Perspectives on: conformational coupling in ion channels: allosteric coupling in ligand-gated ion channels.J Gen Physiol. 2012 Dec;140(6):599-612. doi: 10.1085/jgp.201210844. J Gen Physiol. 2012. PMID: 23183696 Free PMC article. No abstract available.
-
Recent structural and mechanistic insights into endplate acetylcholine receptors.Ann N Y Acad Sci. 2008;1132:53-60. doi: 10.1196/annals.1405.041. Ann N Y Acad Sci. 2008. PMID: 18567853 Free PMC article.
-
Functional anatomy of an allosteric protein.Nat Commun. 2013;4:2984. doi: 10.1038/ncomms3984. Nat Commun. 2013. PMID: 24352193 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
