

The Helicobacter pylori vacuolating toxin inhibits T cell activation by two independent mechanisms
- PMID: 14676300
- PMCID: PMC2194151
- DOI: 10.1084/jem.20030621
The Helicobacter pylori vacuolating toxin inhibits T cell activation by two independent mechanisms
Abstract
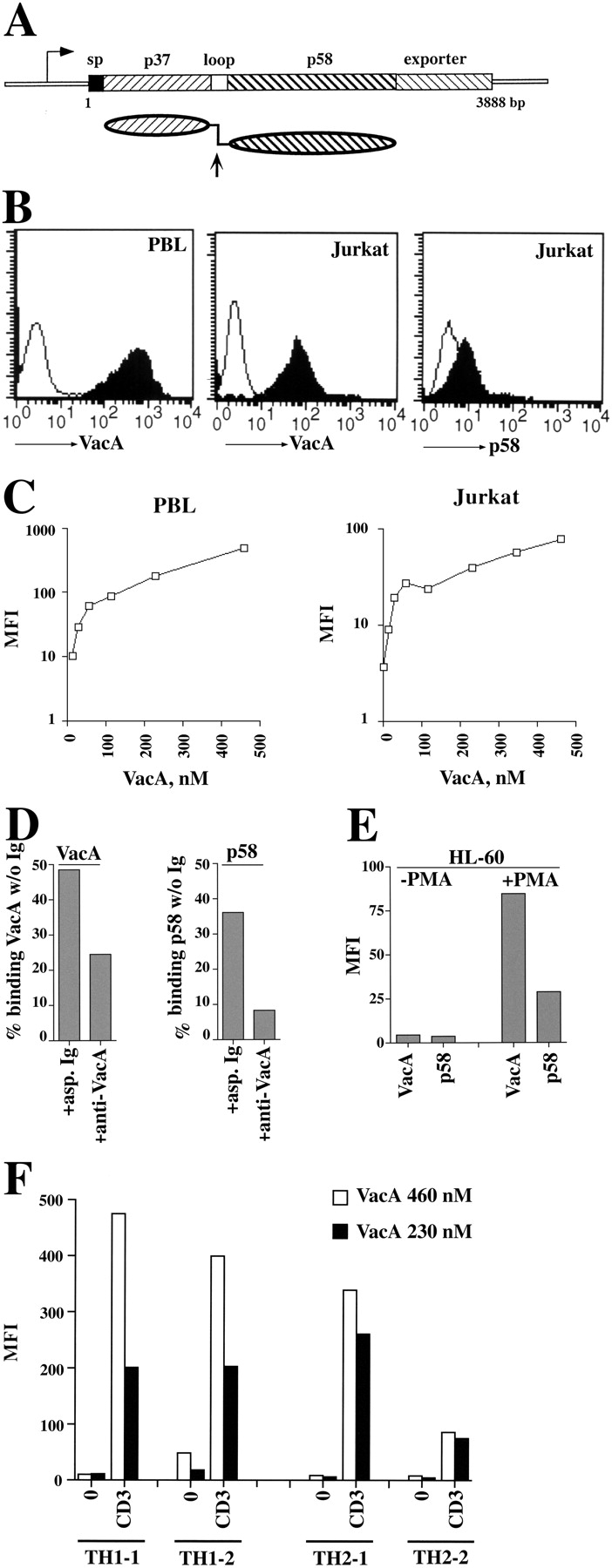
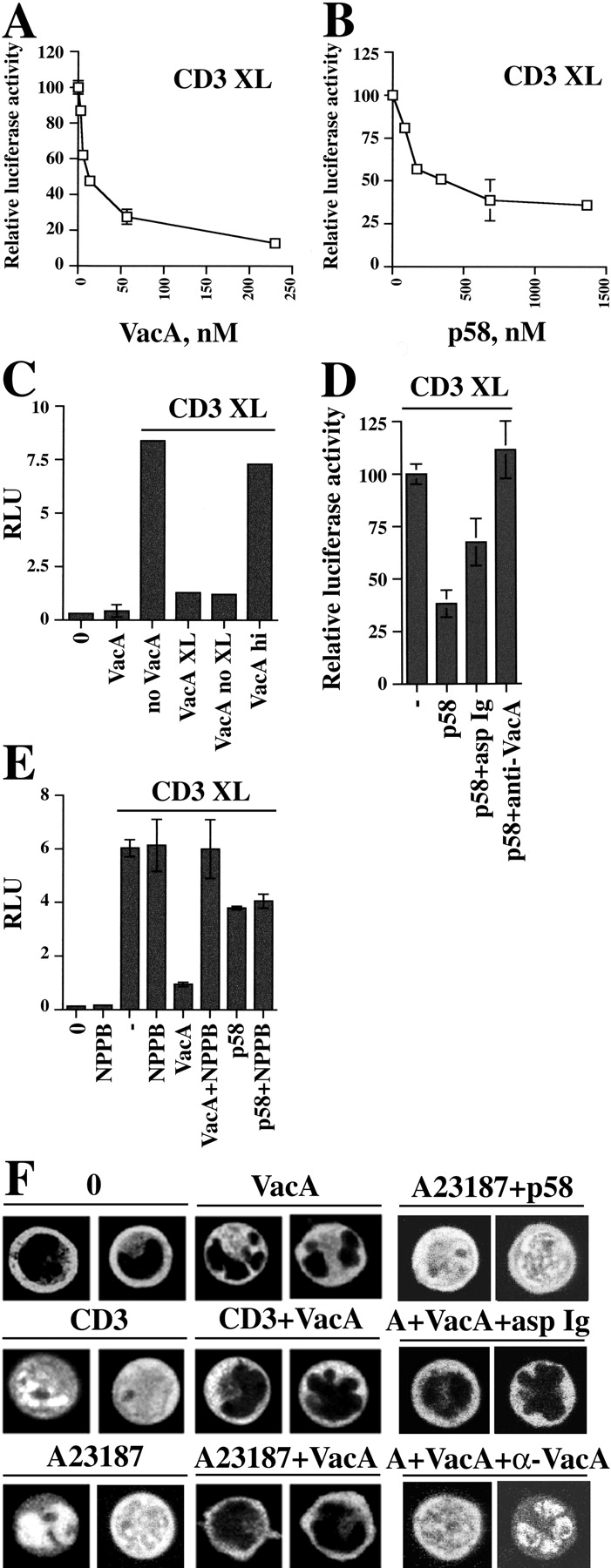
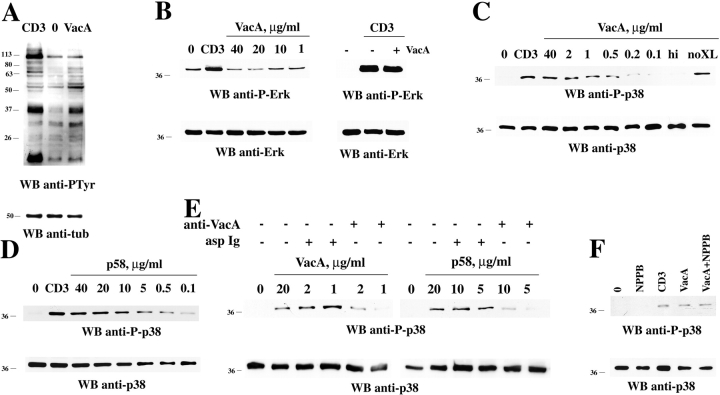
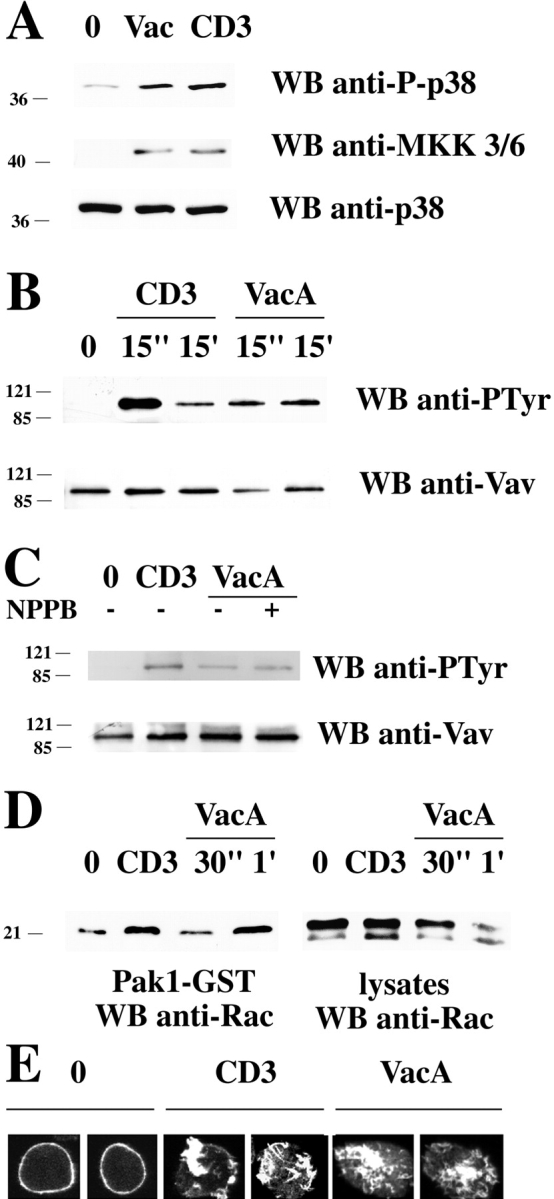
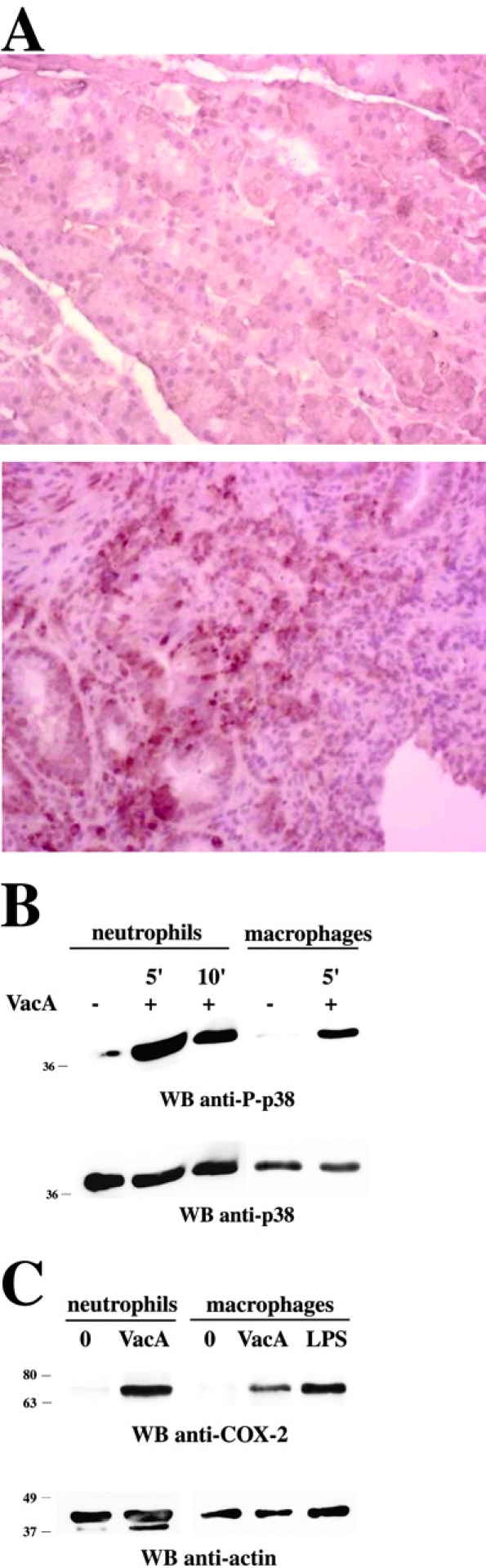
Helicobacter pylori toxin, VacA, damages the gastric epithelium by erosion and loosening of tight junctions. Here we report that VacA also interferes with T cell activation by two different mechanisms. Formation of anion-specific channels by VacA prevents calcium influx from the extracellular milieu. The transcription factor NF-AT thus fails to translocate to the nucleus and activate key cytokine genes. A second, channel-independent mechanism involves activation of intracellular signaling through the mitogen-activated protein kinases MKK3/6 and p38 and the Rac-specific nucleotide exchange factor, Vav. As a consequence of aberrant Rac activation, disordered actin polymerization is stimulated. The resulting defects in T cell activation may help H. pylori to prevent an effective immune response leading to chronic colonization of its gastric niche.
Figures


Comment in
-
Immunosuppressive and proinflammatory activities of the VacA toxin of Helicobacter pylori.J Exp Med. 2003 Dec 15;198(12):1767-71. doi: 10.1084/jem.20031839. J Exp Med. 2003. PMID: 14676291 Free PMC article. No abstract available.
References
-
- Telford, J.L., A. Covacci, P. Ghiara, C. Montecucco, and R. Rappuoli. 1994. Unravelling the pathogenic role of Helicobacter pylori in peptic ulcer: potential new therapies and vaccines. Trends Biotechnol. 12:420–426. - PubMed
-
- Cover, T.L. 1996. The vacuolating cytotoxin of Helicobacter pylori. Mol. Microbiol. 20:241–246. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
