

Abnormal mammary gland development and growth retardation in female mice and MCF7 breast cancer cells lacking androgen receptor
- PMID: 14676301
- PMCID: PMC2194158
- DOI: 10.1084/jem.20031233
Abnormal mammary gland development and growth retardation in female mice and MCF7 breast cancer cells lacking androgen receptor
Abstract
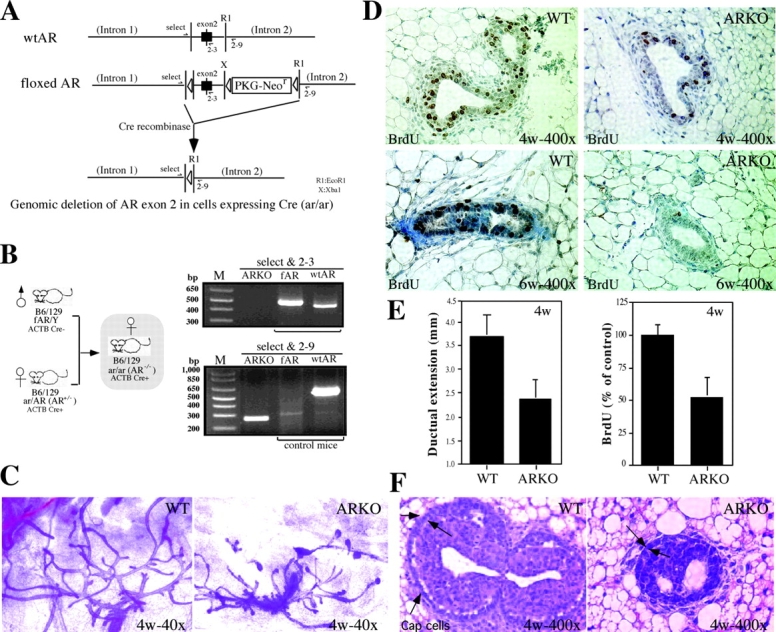
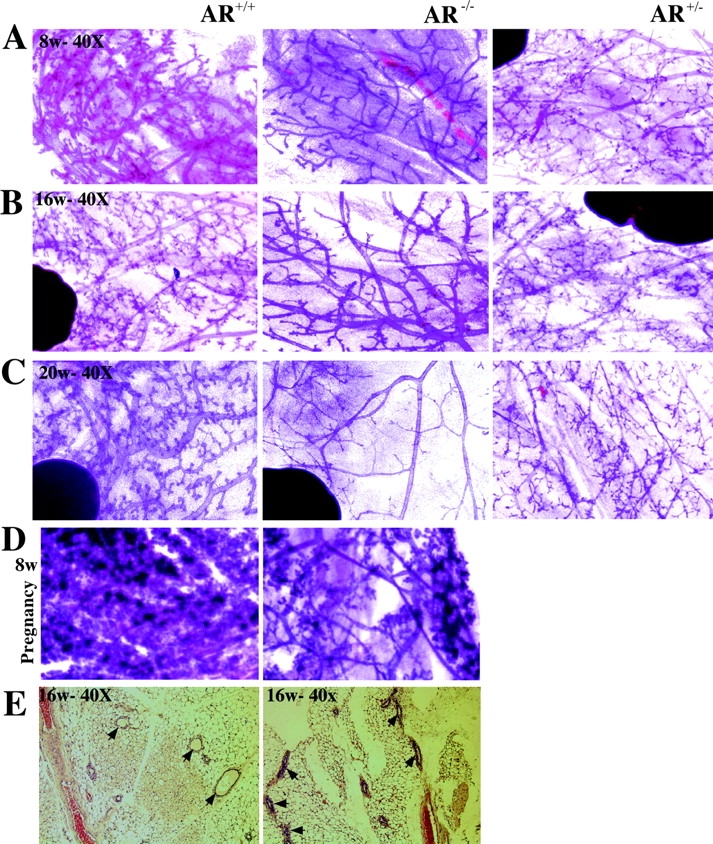
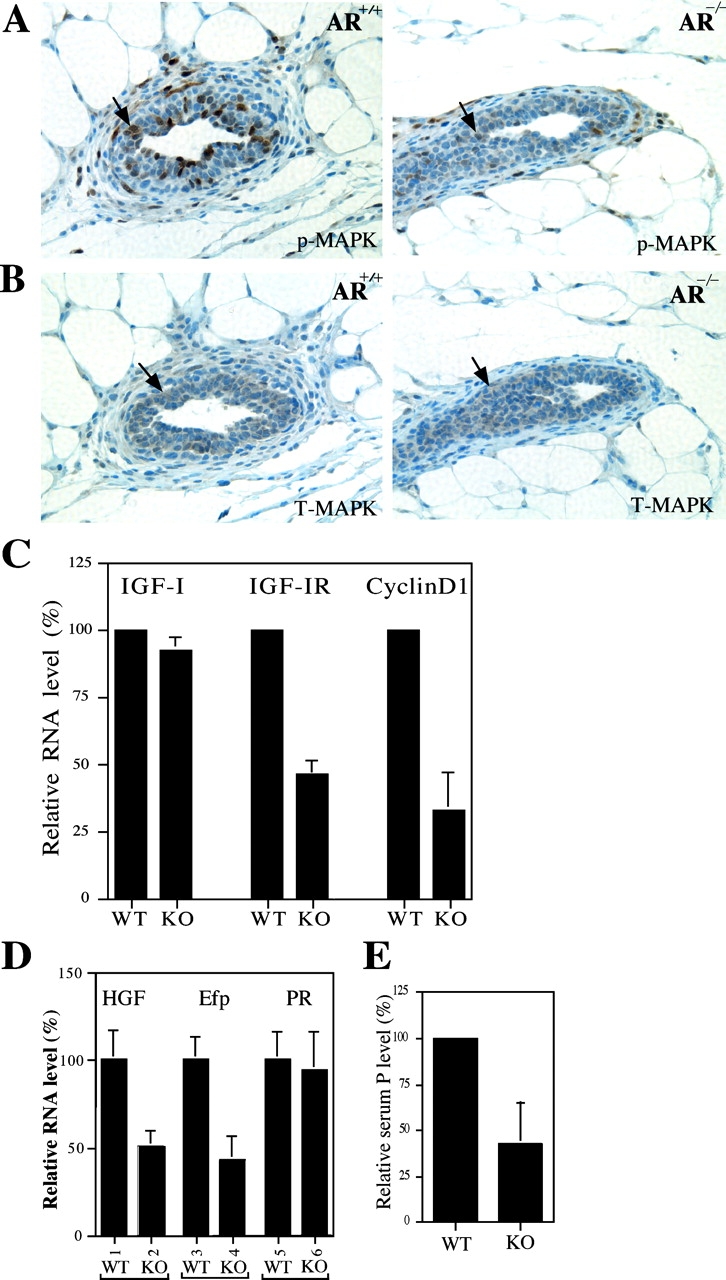
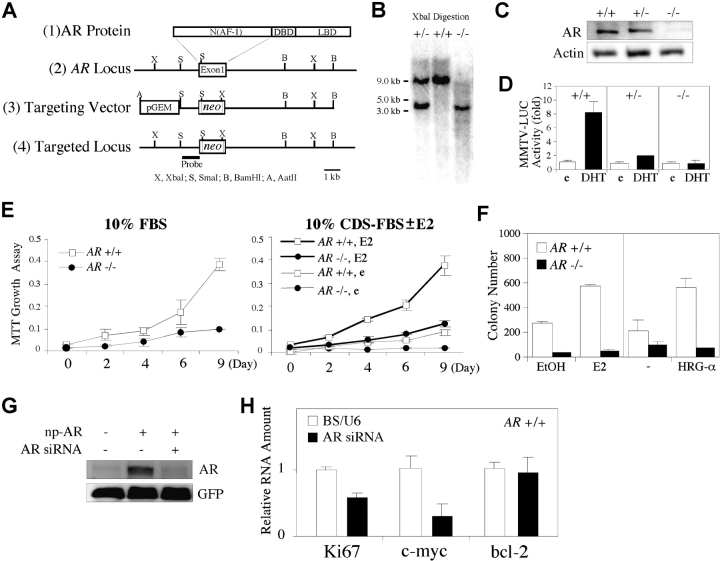
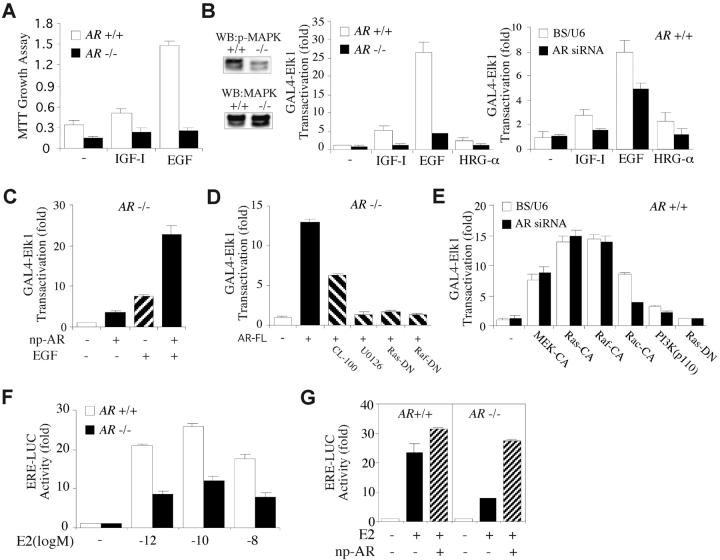
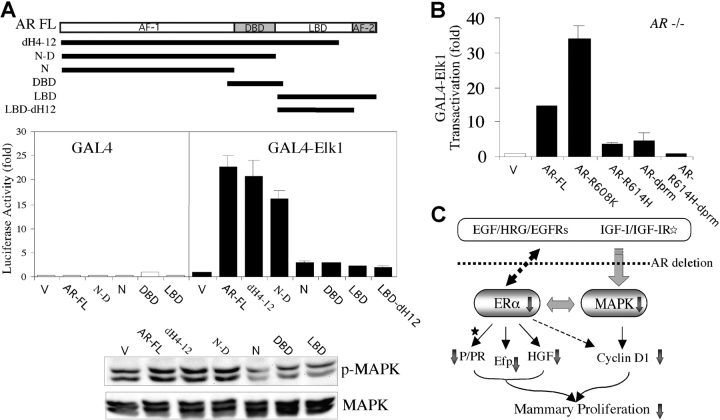
Phenotype analysis of female mice lacking androgen receptor (AR) deficient (AR-/-) indicates that the development of mammary glands is retarded with reduced ductal branching in the prepubertal stages, and fewer Cap cells in the terminal end buds, as well as decreased lobuloalveolar development in adult females, and fewer milk-producing alveoli in the lactating glands. The defective development of AR-/- mammary glands involves the defects of insulin-like growth factor I-insulin-like growth factor I receptor and mitogen-activated protein kinase (MAPK) signals as well as estrogen receptor (ER) activity. Similar growth retardation and defects in growth factor-mediated Ras/Raf/MAPK cascade and ER signaling are also found in AR-/- MCF7 breast cancer cells. The restoration assays show that AR NH2-terminal/DNA-binding domain, but not the ligand-binding domain, is essential for normal MAPK function in MCF7 cells, and an AR mutant (R608K), found in male breast cancer, is associated with the excessive activation of MAPK. Together, our data provide the first in vivo evidence showing that AR-mediated MAPK and ER activation may play important roles for mammary gland development and MCF7 breast cancer cell proliferation.
Figures
References
-
- Chang, C.S., J. Kokontis, and S.T. Liao. 1988. Molecular cloning of human and rat complementary DNA encoding androgen receptors. Science. 240:324–326. - PubMed
-
- McKenna, N.J., and B.W. O'Malley. 2002. Combinatorial control of gene expression by nuclear receptors and coregulators. Cell. 108:465–474. - PubMed
-
- Secreto, G., and B. Zumoff. 1994. Abnormal production of androgens in women with breast cancer. Anticancer Res. 14:2113–2117. - PubMed
-
- Berrino, F., P. Muti, A. Micheli, G. Bolelli, V. Krogh, R. Sciajno, P. Pisani, S. Panico, and G. Secreto. 1996. Serum sex hormone levels after menopause and subsequent breast cancer. J. Natl. Cancer Inst. 88:291–296. - PubMed
-
- Dimitrakakis, C., J. Zhou, and C.A. Bondy. 2002. Androgens and mammary growth and neoplasia. Fertil. Steril. 77(Suppl.):26–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
