

Redox-coupled proton translocation in biological systems: proton shuttling in cytochrome c oxidase
- PMID: 14676323
- PMCID: PMC307604
- DOI: 10.1073/pnas.2432106100
Redox-coupled proton translocation in biological systems: proton shuttling in cytochrome c oxidase
Abstract
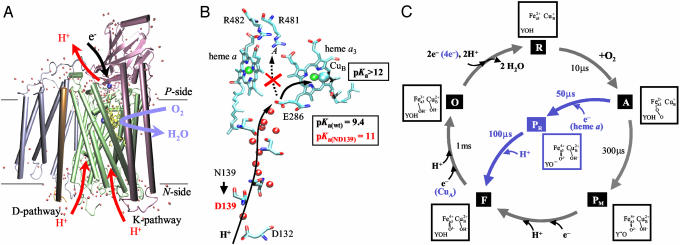
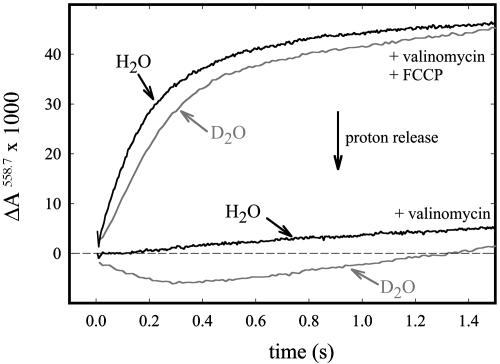
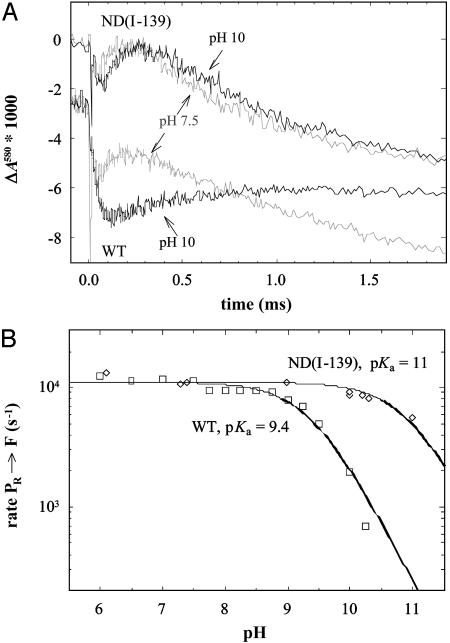
In the respiratory chain free energy is conserved by linking the chemical reduction of dioxygen to the electrogenic translocation of protons across a membrane. Cytochrome c oxidase (CcO) is one of the sites where this linkage occurs. Although intensively studied, the molecular mechanism of proton pumping by this enzyme remains unknown. Here, we present data from an investigation of a mutant CcO from Rhodobacter sphaeroides [Asn-139 --> Asp, ND(I-139)] in which proton pumping is completely uncoupled from the catalytic turnover (i.e., reduction of O2). However, in this mutant CcO, the rate by which O2 is reduced to H2O is even slightly higher than that of the wild-type CcO. The data indicate that the disabling of the proton pump is a result of a perturbation of E(I-286), which is located 20 A from N(I-139) and is an internal proton donor to the catalytic site, located in the membrane-spanning part of CcO. The mutation results in raising the effective pKa of E(I-286) by 1.6 pH units. An explanation of how the mutation uncouples catalytic turnover from proton pumping is offered, which suggests a mechanism by which CcO pumps protons.
Figures
Similar articles
-
Controlled uncoupling and recoupling of proton pumping in cytochrome c oxidase.Proc Natl Acad Sci U S A. 2006 Jan 10;103(2):317-22. doi: 10.1073/pnas.0507734103. Epub 2006 Jan 3. Proc Natl Acad Sci U S A. 2006. PMID: 16407159 Free PMC article.
-
G204D, a mutation that blocks the proton-conducting D-channel of the aa3-type cytochrome c oxidase from Rhodobacter sphaeroides.Biochemistry. 2005 Sep 27;44(38):12767-74. doi: 10.1021/bi051141m. Biochemistry. 2005. PMID: 16171391
-
Variable proton-pumping stoichiometry in structural variants of cytochrome c oxidase.Biochim Biophys Acta. 2010 Jun-Jul;1797(6-7):710-23. doi: 10.1016/j.bbabio.2010.02.020. Epub 2010 Feb 23. Biochim Biophys Acta. 2010. PMID: 20184858 Review.
-
Aspartate-132 in cytochrome c oxidase from Rhodobacter sphaeroides is involved in a two-step proton transfer during oxo-ferryl formation.Biochemistry. 1999 May 25;38(21):6826-33. doi: 10.1021/bi982865j. Biochemistry. 1999. PMID: 10346904
-
Molecular mechanism of proton translocation by cytochrome c oxidase.Antioxid Redox Signal. 2008 Jan;10(1):1-29. doi: 10.1089/ars.2007.1705. Antioxid Redox Signal. 2008. PMID: 17949262 Review.
Cited by
-
Controlled uncoupling and recoupling of proton pumping in cytochrome c oxidase.Proc Natl Acad Sci U S A. 2006 Jan 10;103(2):317-22. doi: 10.1073/pnas.0507734103. Epub 2006 Jan 3. Proc Natl Acad Sci U S A. 2006. PMID: 16407159 Free PMC article.
-
Multiscale simulations reveal key features of the proton-pumping mechanism in cytochrome c oxidase.Proc Natl Acad Sci U S A. 2016 Jul 5;113(27):7420-5. doi: 10.1073/pnas.1601982113. Epub 2016 Jun 23. Proc Natl Acad Sci U S A. 2016. PMID: 27339133 Free PMC article.
-
Decoupling mutations in the D-channel of the aa(3)-type cytochrome c oxidase from Rhodobacter sphaeroides suggest that a continuous hydrogen-bonded chain of waters is essential for proton pumping.Biochemistry. 2010 Jun 1;49(21):4476-82. doi: 10.1021/bi100344x. Biochemistry. 2010. PMID: 20441187 Free PMC article.
-
Alternative initial proton acceptors for the D pathway of Rhodobacter sphaeroides cytochrome c oxidase.Biochemistry. 2011 Apr 12;50(14):2820-8. doi: 10.1021/bi102002v. Epub 2011 Mar 21. Biochemistry. 2011. PMID: 21344856 Free PMC article.
-
Proton-pumping mechanism of cytochrome c oxidase: a kinetic master-equation approach.Biochim Biophys Acta. 2012 Apr;1817(4):526-36. doi: 10.1016/j.bbabio.2011.09.004. Epub 2011 Sep 16. Biochim Biophys Acta. 2012. PMID: 21946020 Free PMC article. Review.
References
-
- Ferguson-Miller, S. & Babcock, G. T. (1996) Chem. Rev. 96, 2889–2907. - PubMed
-
- Babcock, G. T. & Wikström, M. (1992) Nature 356, 301–309. - PubMed
-
- Zaslavsky, D. & Gennis, R. B. (2000) Biochim. Biophys. Acta 1458, 164–179. - PubMed
-
- Tsukihara, T., Aoyama, H., Yamashita, E., Tomizaki, T., Yamaguchi, H., Shinzawa-Itoh, K., Nakashima, R., Yaono, R. & Yoshikawa, S. (1996) Science 272, 1136–1144. - PubMed
-
- Iwata, S., Ostermeier, C., Ludwig, B. & Michel, H. (1995) Nature 376, 660–669. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
