

Antisense-induced multiexon skipping for Duchenne muscular dystrophy makes more sense
- PMID: 14681829
- PMCID: PMC1181915
- DOI: 10.1086/381039
Antisense-induced multiexon skipping for Duchenne muscular dystrophy makes more sense
Abstract
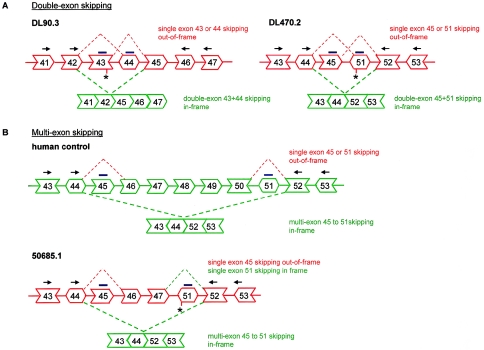
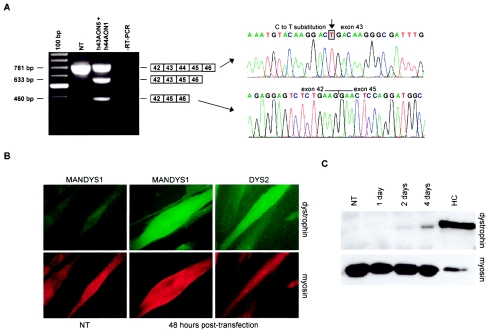
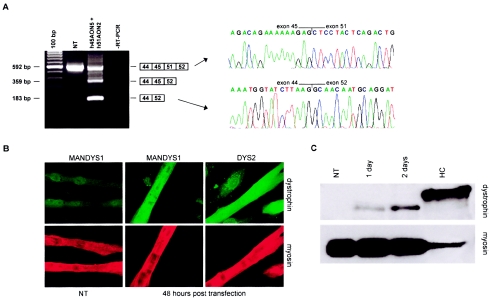
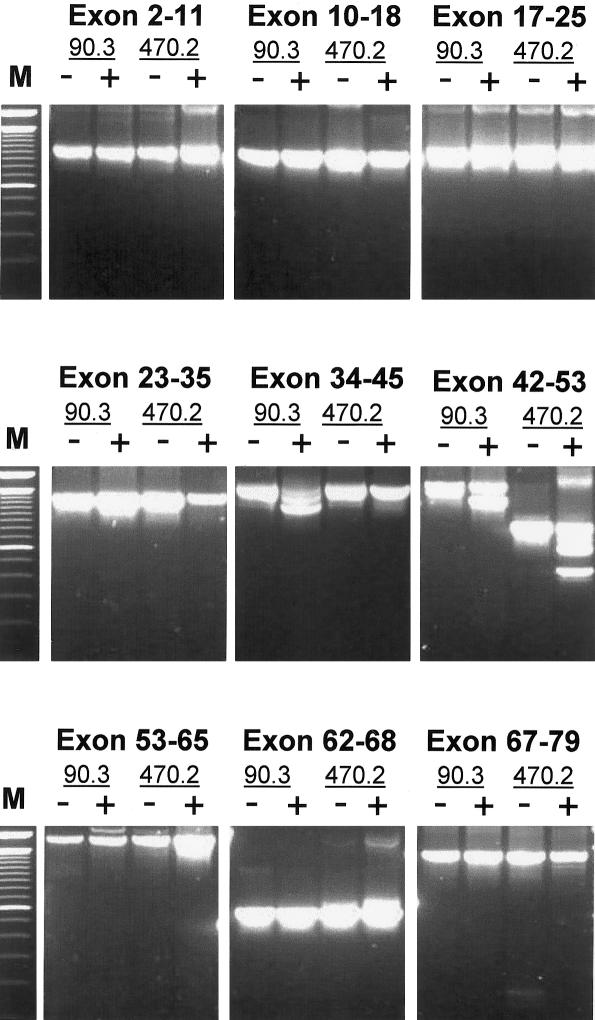
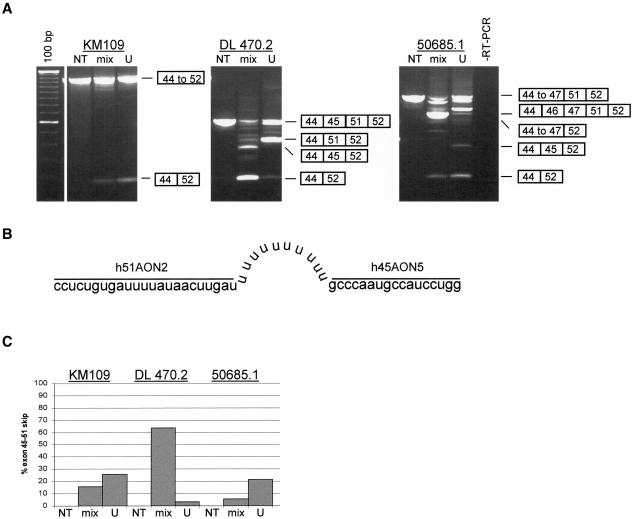
Dystrophin deficiency, which leads to severe and progressive muscle degeneration in patients with Duchenne muscular dystrophy (DMD), is caused by frameshifting mutations in the dystrophin gene. A relatively new therapeutic strategy is based on antisense oligonucleotides (AONs) that induce the specific skipping of a single exon, such that the reading frame is restored. This allows the synthesis of a largely functional dystrophin, associated with a milder Becker muscular dystrophy phenotype. We have previously successfully targeted 20 different DMD exons that would, theoretically, be beneficial for >75% of all patients. To further enlarge this proportion, we here studied the feasibility of double and multiexon skipping. Using a combination of AONs, double skipping of exon 43 and 44 was induced, and dystrophin synthesis was restored in myotubes from one patient affected by a nonsense mutation in exon 43. For another patient, with an exon 46-50 deletion, the therapeutic double skipping of exon 45 and 51 was achieved. Remarkably, in control myotubes, the latter combination of AONs caused the skipping of the entire stretch of exons from 45 through 51. This in-frame multiexon skipping would be therapeutic for a series of patients carrying different DMD-causing mutations. In fact, we here demonstrate its feasibility in myotubes from a patient with an exon 48-50 deletion. The application of multiexon skipping may provide a more uniform methodology for a larger group of patients with DMD.
Figures
References
Electronic-Database Information
-
- Online Mendelian Inheritance in Man (OMIM), http://www.ncbi.nlm.nih.gov/Omim/ (for DMD and BMD)
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
