

Lecithin-retinol acyltransferase is essential for accumulation of all-trans-retinyl esters in the eye and in the liver
- PMID: 14684738
- PMCID: PMC1351249
- DOI: 10.1074/jbc.M312410200
Lecithin-retinol acyltransferase is essential for accumulation of all-trans-retinyl esters in the eye and in the liver
Abstract
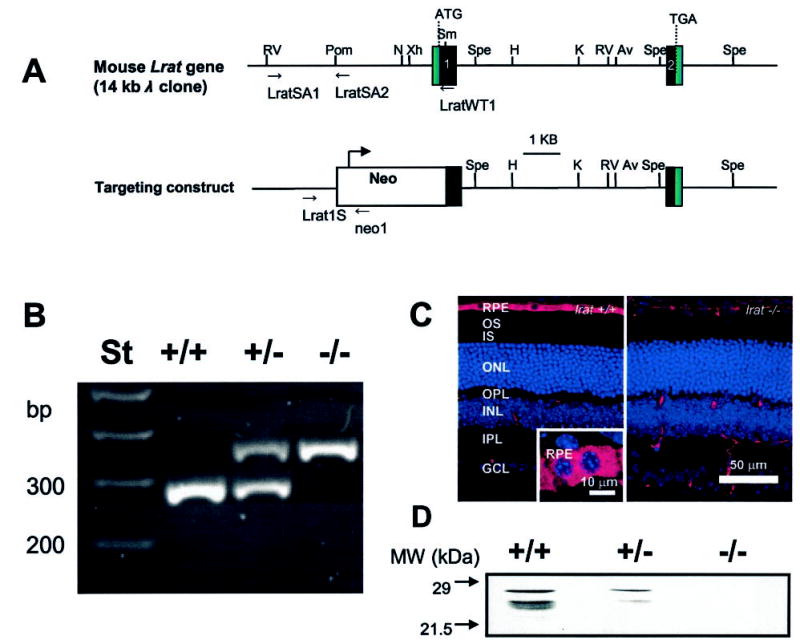
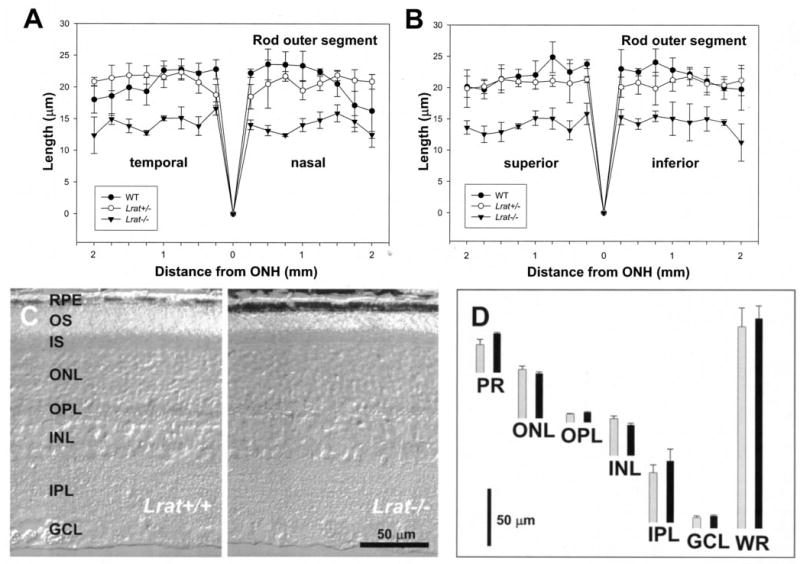
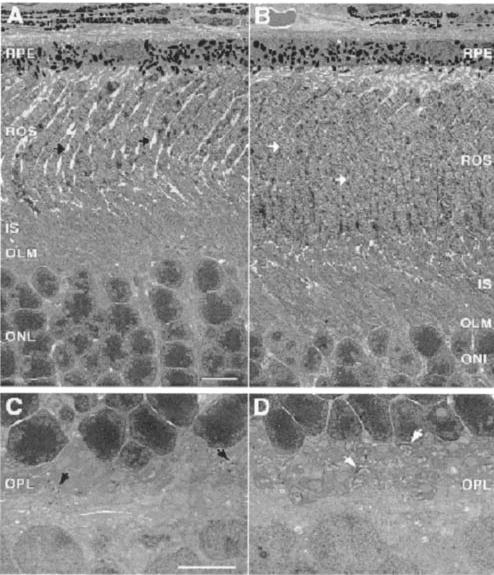
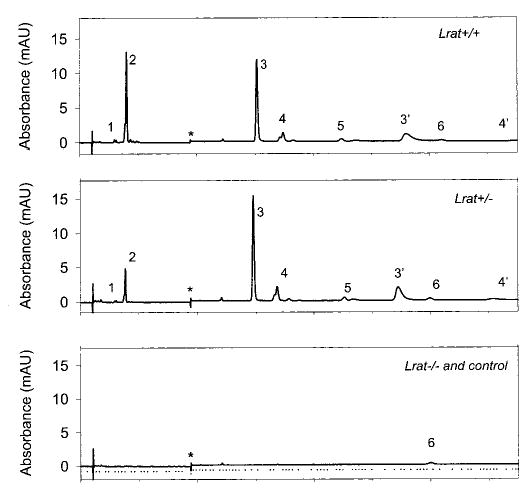
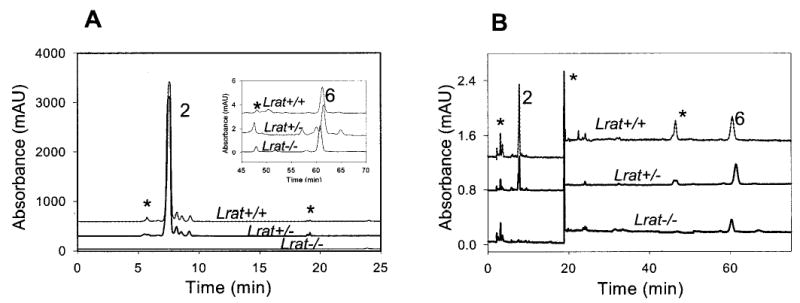
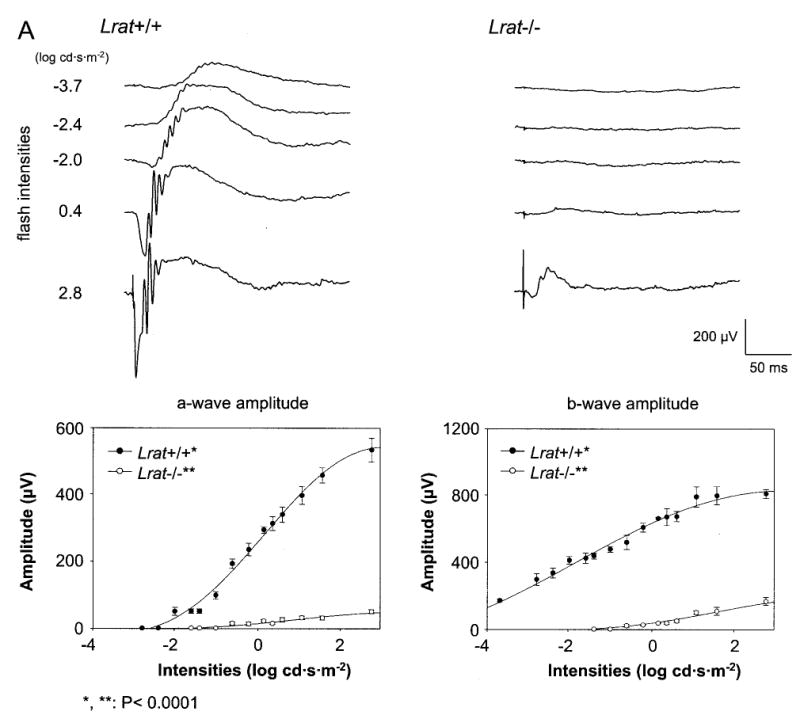
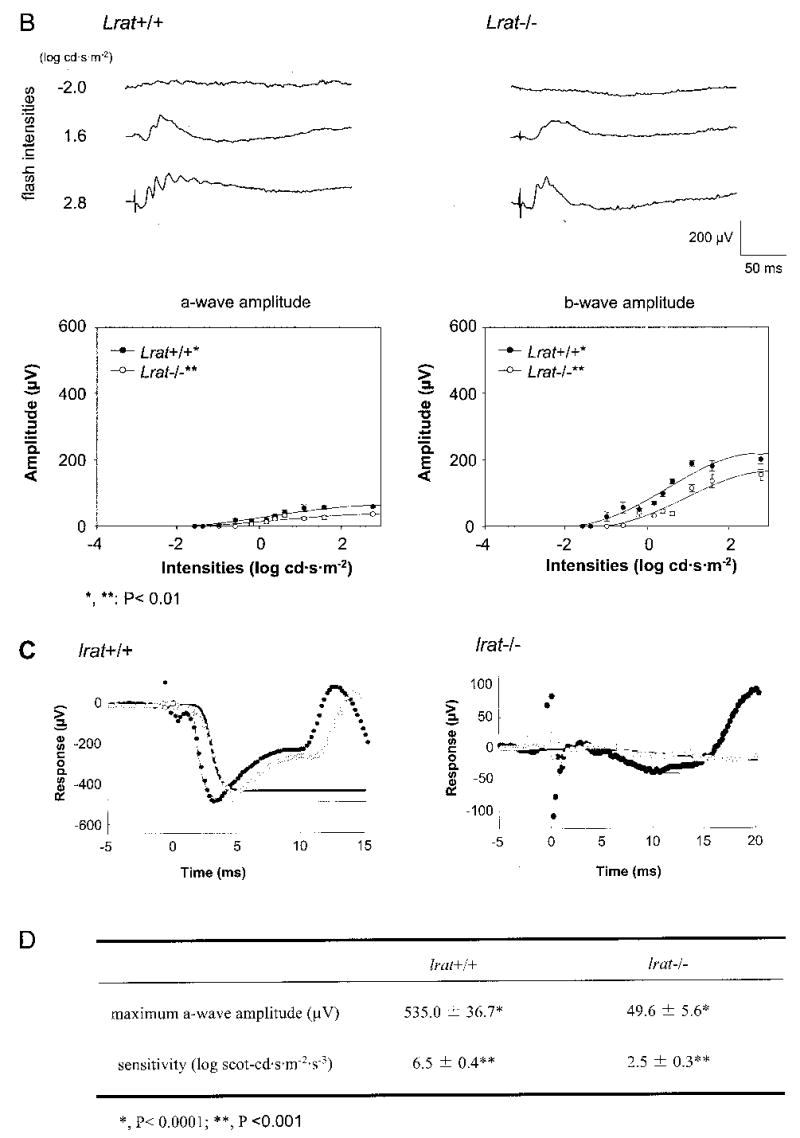
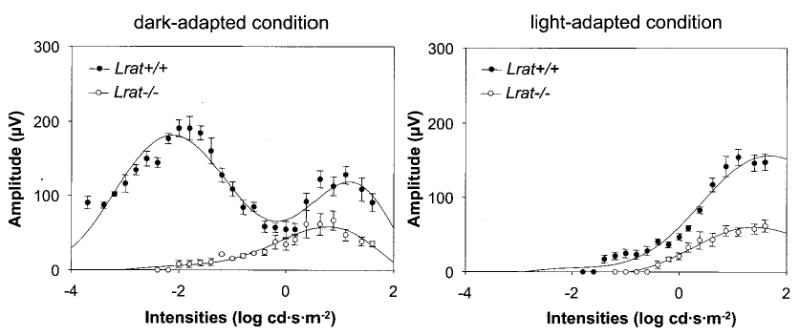
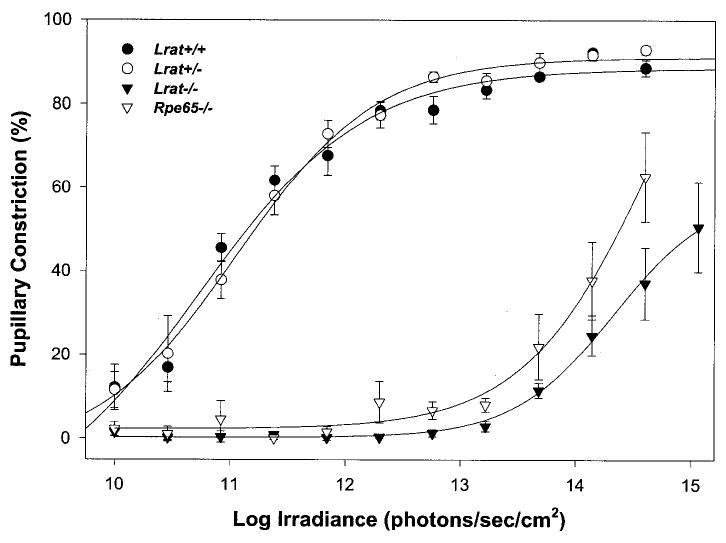
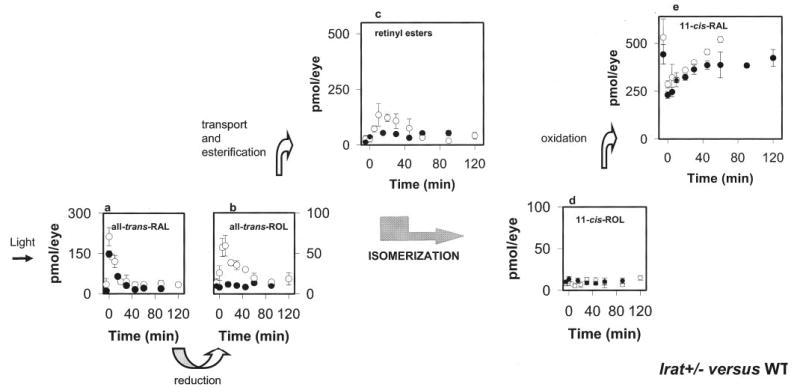
Lecithin-retinol acyltransferase (LRAT), an enzyme present mainly in the retinal pigmented epithelial cells and liver, converts all-trans-retinol into all-trans-retinyl esters. In the retinal pigmented epithelium, LRAT plays a key role in the retinoid cycle, a two-cell recycling system that replenishes the 11-cis-retinal chromophore of rhodopsin and cone pigments. We disrupted mouse Lrat gene expression by targeted recombination and generated a homozygous Lrat knock-out (Lrat-/-) mouse. Despite the expression of LRAT in multiple tissues, the Lrat-/- mouse develops normally. The histological analysis and electron microscopy of the retina for 6-8-week-old Lrat-/- mice revealed that the rod outer segments are approximately 35% shorter than those of Lrat+/+ mice, whereas other neuronal layers appear normal. Lrat-/- mice have trace levels of all-trans-retinyl esters in the liver, lung, eye, and blood, whereas the circulating all-trans-retinol is reduced only slightly. Scotopic and photopic electroretinograms as well as pupillary constriction analyses revealed that rod and cone visual functions are severely attenuated at an early age. We conclude that Lrat-/- mice may serve as an animal model with early onset severe retinal dystrophy and severe retinyl ester deprivation.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
