

Maize mutants lacking chloroplast FtsY exhibit pleiotropic defects in the biogenesis of thylakoid membranes
- PMID: 14688289
- PMCID: PMC301405
- DOI: 10.1105/tpc.014787
Maize mutants lacking chloroplast FtsY exhibit pleiotropic defects in the biogenesis of thylakoid membranes
Abstract
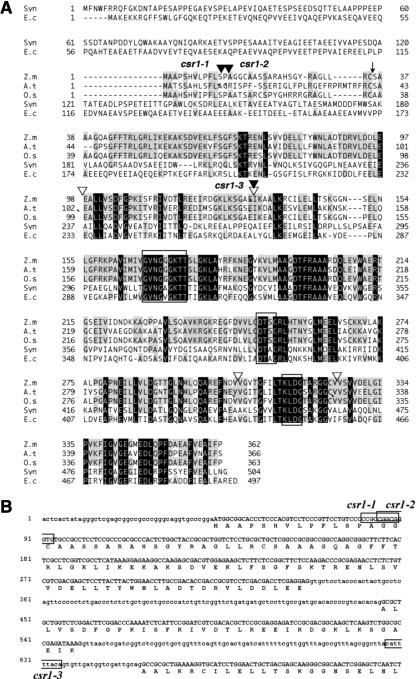
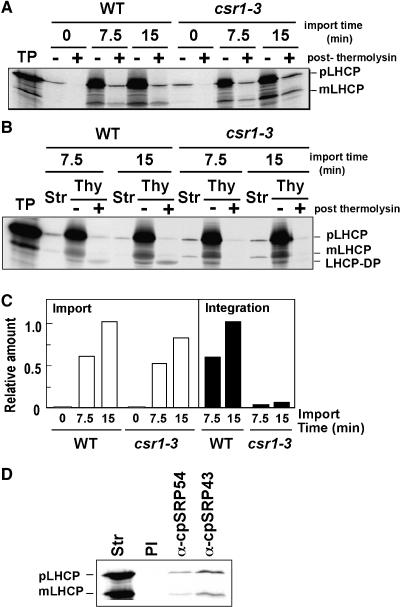
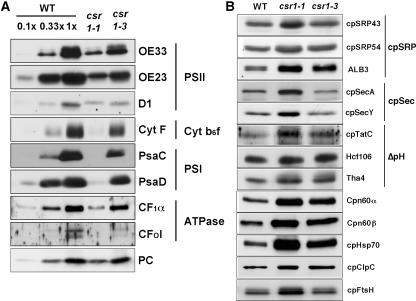
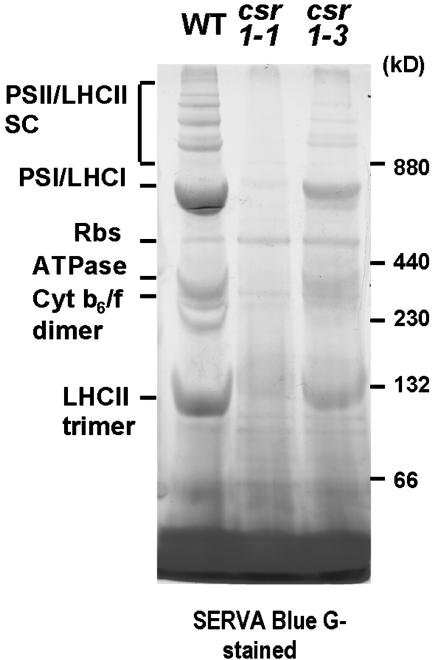
A chloroplast signal recognition particle (SRP) that is related to the SRP involved in secretion in bacteria and eukaryotic cells is used for the insertion of light-harvesting chlorophyll proteins (LHCPs) into the thylakoid membranes. A conserved component of the SRP mechanism is a membrane-bound SRP receptor, denoted FtsY in bacteria. Plant genomes encode FtsY homologs that are targeted to the chloroplast (cpFtsY). To investigate the in vivo roles of cpFtsY, we characterized maize cpFtsY and maize mutants having a Mu transposon insertion in the corresponding gene (chloroplast SRP receptor1, or csr1). Maize cpFtsY accumulates to much higher levels in leaf tissue than in roots and stems. Interestingly, it is present at similar levels in etiolated and green leaf tissue and was found to bind the prolamellar bodies of etioplasts. A null cpFtsY mutant, csr1-1, showed a substantial loss of leaf chlorophyll, whereas a "leaky" allele, csr1-3, conditioned a more moderate chlorophyll deficiency. Both alleles caused the loss of various LHCPs and the thylakoid-bound photosynthetic enzyme complexes and were seedling lethal. By contrast, levels of the membrane-bound components of the thylakoid protein transport machineries were not altered. The thylakoid membranes in csr1-1 chloroplasts were unstacked and reduced in abundance, but the prolamellar bodies in mutant etioplasts appeared normal. These results demonstrate the essentiality of cpFtsY for the biogenesis not only of the LHCPs but also for the assembly of the other membrane-bound components of the photosynthetic apparatus.
Figures




References
-
- Allen, J.F., and Forsberg, J. (2001). Molecular recognition in thylakoid structure and function. Trends Plant Sci. 6, 317–326. - PubMed
-
- Aronsson, H., Sundqvist, C., and Dahlin, C. (2003). POR: Import and membrane association of a key element in chloroplast development. Physiol. Plant. 118, 1–9. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
