

A role for cytoplasmic dynein and LIS1 in directed cell movement
- PMID: 14691133
- PMCID: PMC2173723
- DOI: 10.1083/jcb.200310097
A role for cytoplasmic dynein and LIS1 in directed cell movement
Abstract
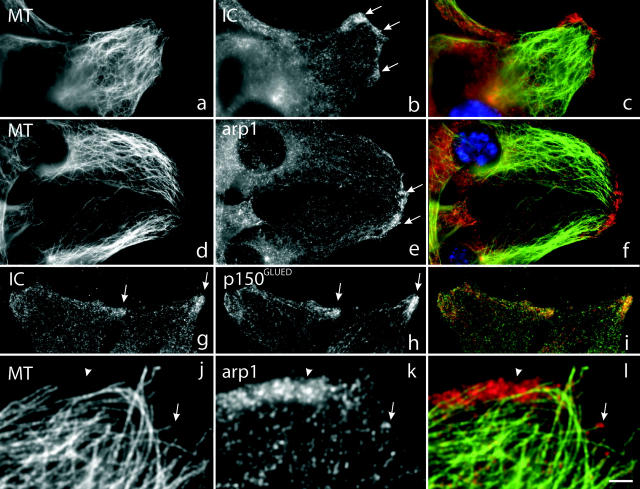
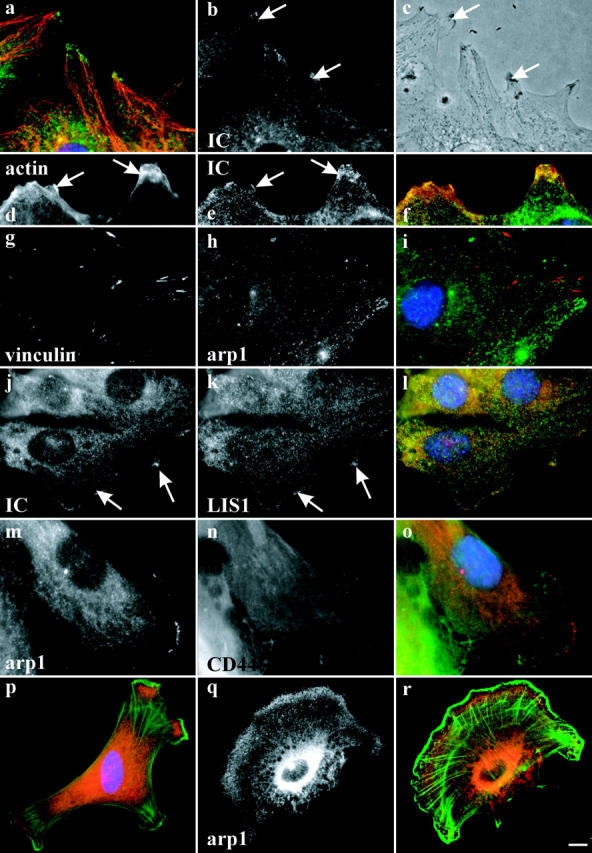
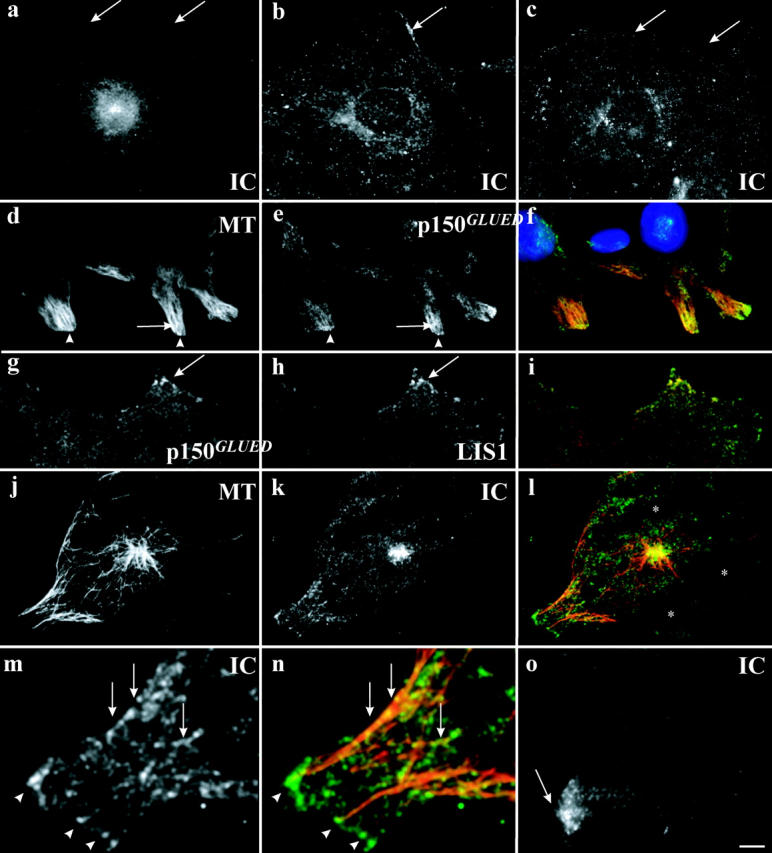
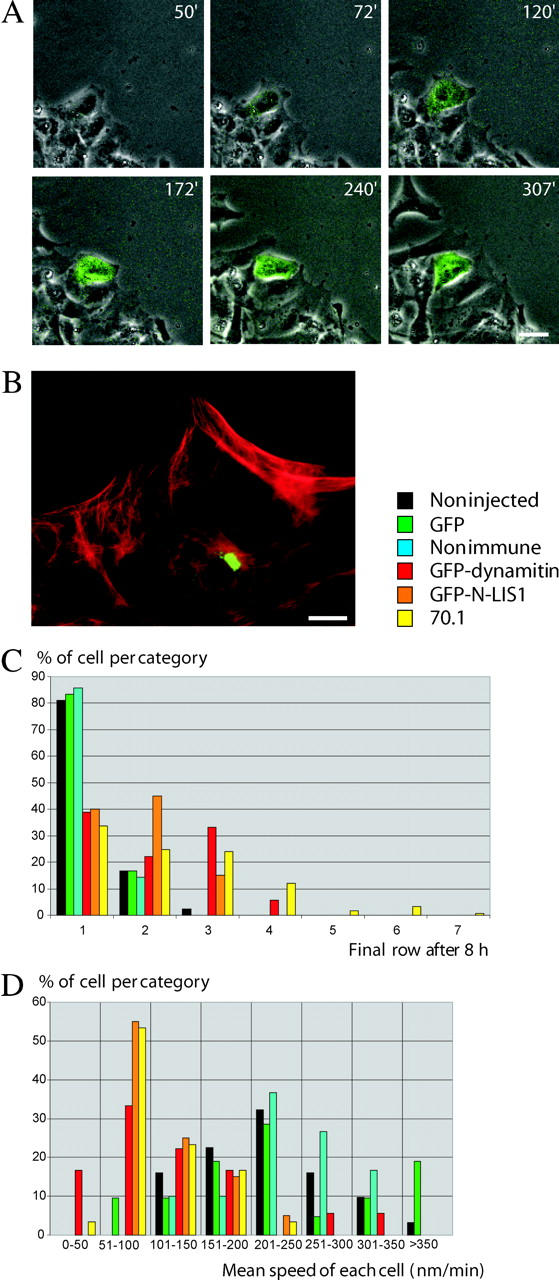
Cytoplasmic dynein has been implicated in numerous aspects of intracellular movement. We recently found dynein inhibitors to interfere with the reorientation of the microtubule cytoskeleton during healing of wounded NIH3T3 cell monolayers. We now find that dynein and its regulators dynactin and LIS1 localize to the leading cell cortex during this process. In the presence of serum, bright diffuse staining was observed in regions of active ruffling. This pattern was abolished by cytochalasin D, and was not observed in cells treated with lysophosphatidic acid, conditions which allow microtubule reorientation but not forward cell movement. Under the same conditions, using total internal reflection fluorescence microscopy, clear punctate dynein/dynactin containing structures were observed along the sides and at the tips of microtubules at the leading edge. Overexpression of dominant negative dynactin and LIS1 cDNAs or injection of antidynein antibody interfered with the rate of cell migration. Together, these results implicate a leading edge cortical pool of dynein in both early and persistent steps in directed cell movement.
Figures
References
-
- Busson, S., D. Dujardin, A. Moreau, J. Dompierre, and J.R. De Mey. 1998. Dynein and dynactin are localized to astral microtubules and at cortical sites in mitotic epithelial cells. Curr. Biol. 8:541–544. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
