

N-linked glycosylation of dipeptidyl peptidase IV (CD26): effects on enzyme activity, homodimer formation, and adenosine deaminase binding
- PMID: 14691230
- PMCID: PMC2286525
- DOI: 10.1110/ps.03352504
N-linked glycosylation of dipeptidyl peptidase IV (CD26): effects on enzyme activity, homodimer formation, and adenosine deaminase binding
Abstract
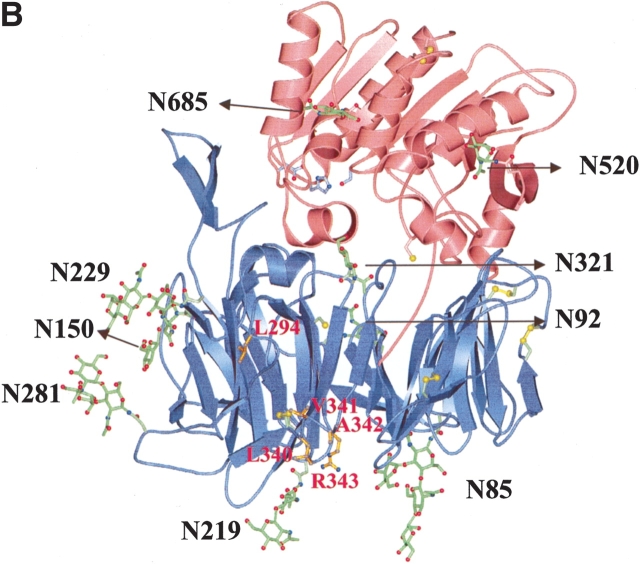
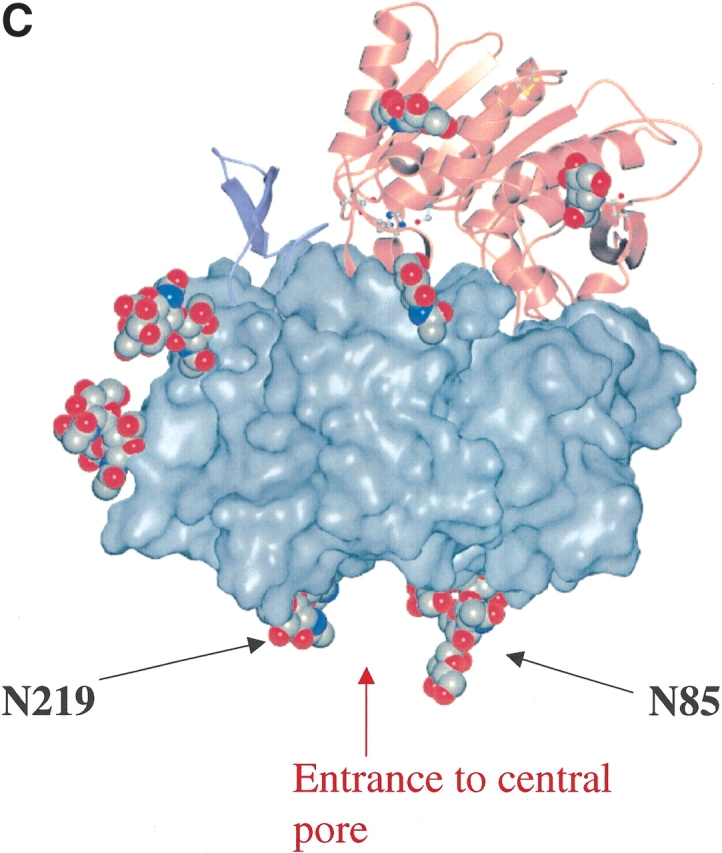
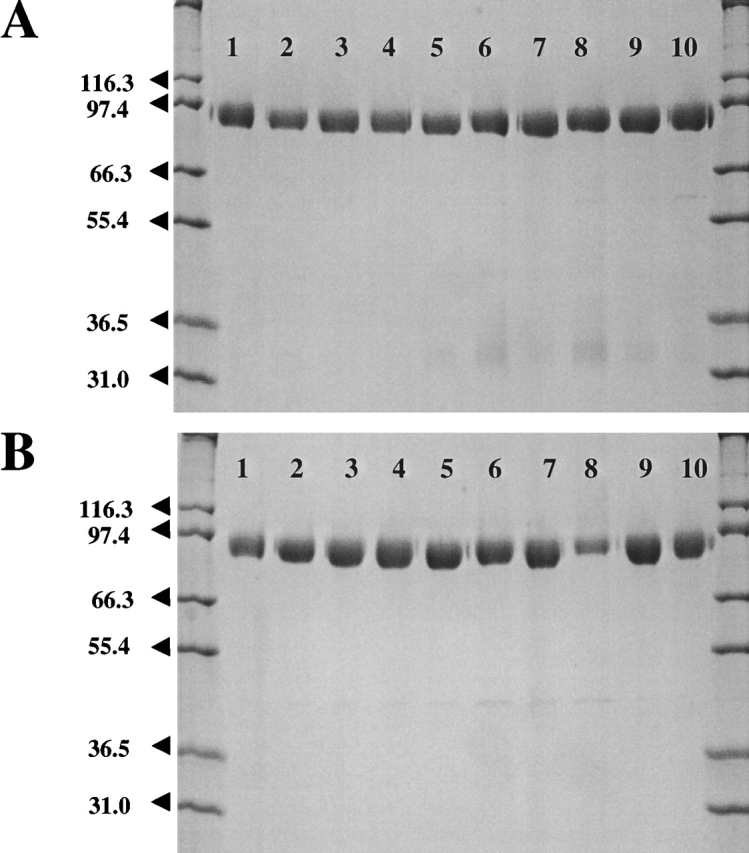
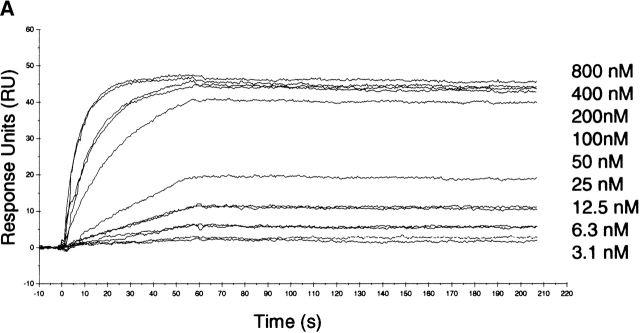
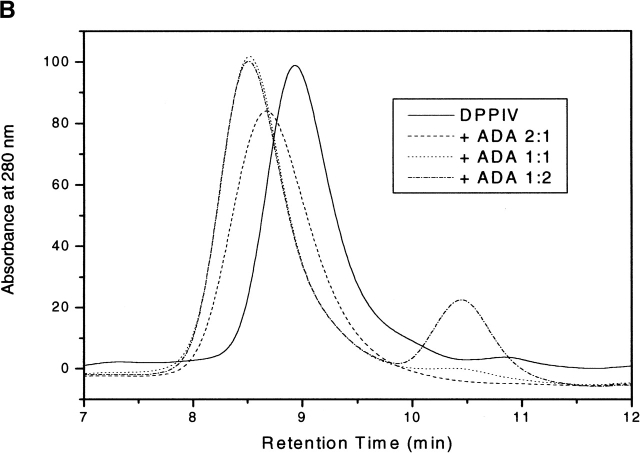
The type II transmembrane serine protease dipeptidyl peptidase IV (DPPIV), also known as CD26 or adenosine deaminase binding protein, is a major regulator of various physiological processes, including immune, inflammatory, nervous, and endocrine functions. It has been generally accepted that glycosylation of DPPIV and of other transmembrane dipeptidyl peptidases is a prerequisite for enzyme activity and correct protein folding. Crystallographic studies on DPPIV reveal clear N-linked glycosylation of nine Asn residues in DPPIV. However, the importance of each glycosylation site on physiologically relevant reactions such as dipeptide cleavage, dimer formation, and adenosine deaminase (ADA) binding remains obscure. Individual Asn-->Ala point mutants were introduced at the nine glycosylation sites in the extracellular domain of DPPIV (residues 39-766). Crystallographic and biochemical data demonstrate that N-linked glycosylation of DPPIV does not contribute significantly to its peptidase activity. The kinetic parameters of dipeptidyl peptidase cleavage of wild-type DPPIV and the N-glycosylation site mutants were determined by using Ala-Pro-AFC and Gly-Pro-pNA as substrates and varied by <50%. DPPIV is active as a homodimer. Size-exclusion chromatographic analysis showed that the glycosylation site mutants do not affect dimerization. ADA binds to the highly glycosylated beta-propeller domain of DPPIV, but the impact of glycosylation on binding had not previously been determined. Our studies indicate that glycosylation of DPPIV is not required for ADA binding. Taken together, these data indicate that in contrast to the generally accepted view, glycosylation of DPPIV is not a prerequisite for catalysis, dimerization, or ADA binding.
Figures

Similar articles
-
Binding to human dipeptidyl peptidase IV by adenosine deaminase and antibodies that inhibit ligand binding involves overlapping, discontinuous sites on a predicted beta propeller domain.Eur J Biochem. 1999 Dec;266(3):798-810. doi: 10.1046/j.1432-1327.1999.00902.x. Eur J Biochem. 1999. PMID: 10583373
-
Crystal structure of CD26/dipeptidyl-peptidase IV in complex with adenosine deaminase reveals a highly amphiphilic interface.J Biol Chem. 2004 Oct 8;279(41):43330-5. doi: 10.1074/jbc.M405001200. Epub 2004 Jun 22. J Biol Chem. 2004. PMID: 15213224
-
Influence of dipeptidyl peptidase IV on enzymatic properties of adenosine deaminase.Acta Biochim Pol. 2006;53(3):539-46. Epub 2006 Aug 21. Acta Biochim Pol. 2006. PMID: 16929383
-
Molecular mechanism and structural basis of interactions of dipeptidyl peptidase IV with adenosine deaminase and human immunodeficiency virus type-1 transcription transactivator.Eur J Cell Biol. 2012 Apr;91(4):265-73. doi: 10.1016/j.ejcb.2011.06.001. Epub 2011 Sep 28. Eur J Cell Biol. 2012. PMID: 21856036 Review.
-
Peptide truncation by dipeptidyl peptidase IV: a new pathway for drug discovery?Verh K Acad Geneeskd Belg. 2001;63(1):5-32; discussion 32-3. Verh K Acad Geneeskd Belg. 2001. PMID: 11284388 Review.
Cited by
-
Structural Similarities, in Relation with the Cross-Reactivity, of Hymenoptera Allergenic Dipeptidyl Peptidases IV-An Overall Comparison Including a New Dipeptidyl Peptidase IV Sequence from Vespa velutina.Toxins (Basel). 2023 Nov 14;15(11):656. doi: 10.3390/toxins15110656. Toxins (Basel). 2023. PMID: 37999519 Free PMC article.
-
Genetic Insights into the Middle East Respiratory Syndrome Coronavirus Infection among Saudi People.Vaccines (Basel). 2021 Oct 17;9(10):1193. doi: 10.3390/vaccines9101193. Vaccines (Basel). 2021. PMID: 34696302 Free PMC article.
-
Dipeptidyl Peptidase-4 and Adolescent Idiopathic Scoliosis: Expression in Osteoblasts.Sci Rep. 2017 Jun 9;7(1):3173. doi: 10.1038/s41598-017-03310-x. Sci Rep. 2017. PMID: 28600546 Free PMC article.
-
N-glycosylation in Spodoptera frugiperda (Lepidoptera: Noctuidae) midgut membrane-bound glycoproteins.Comp Biochem Physiol B Biochem Mol Biol. 2020 Aug-Sep;246-247:110464. doi: 10.1016/j.cbpb.2020.110464. Epub 2020 Jun 14. Comp Biochem Physiol B Biochem Mol Biol. 2020. PMID: 32553552 Free PMC article.
-
Histoplasma capsulatum encodes a dipeptidyl peptidase active against the mammalian immunoregulatory peptide, substance P.PLoS One. 2009;4(4):e5281. doi: 10.1371/journal.pone.0005281. Epub 2009 Apr 22. PLoS One. 2009. PMID: 19384411 Free PMC article.
References
-
- Abbott, C.A., Baker, E., Sutherland, G.R., and McCaughan, G.W. 1994. Genomic organization, exact localization, and tissue expression of the human CD26 (dipeptidyl peptidase IV) gene. Immunogenetics 40 331–338. - PubMed
-
- Abbott, C.A., McCaughan, G.W., Levy, M.T., Church, W.B., and Gorell, M.D. 1999. Binding to human dipeptidyl peptidase IV by adenosine deaminase and antibodies that inhibit ligand binding involves overlapping, discontinuous sites on a predicted β propeller domain. Eur. J. Biochem. 266 798–810. - PubMed
-
- Abbott, C.A., Yu, D., McCaughan, G.W., and Gorrell, M.D. 2000a. Post-proline-cleaving peptidases having DP IV like enzyme activity: Post-proline peptidases. Exp. Med. Biol. 477 103–109. - PubMed
-
- Abbott, C.A., Yu, D.M., Woollatt, E., Sutherland, G.R., McCaughan, G.W., and Gorrell, M.D. 2000b. Cloning, expression and chromosomal localization of a novel human dipeptidyl peptidase (DPP) IV homolog, DPP8. Eur. J. Biochem. 267 6140–6150. - PubMed
-
- Ahren, B., Simonsson, E., Larsson, H., Landin-Olsson, M., Torgeirsson, H., Jansson, P.A., Sandqvist, M., Bavenholm, P., Efendic, S., Eriksson, J.W., et al. 2002. Inhibition of dipeptidyl peptidase IV improves metabolic control over a 4-week study period in type 2 diabetes. Diabetes Care 25 869–875. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
