

Mutant human cells with constitutive activation of NF-kappaB
- PMID: 14691254
- PMCID: PMC314161
- DOI: 10.1073/pnas.0306812101
Mutant human cells with constitutive activation of NF-kappaB
Abstract
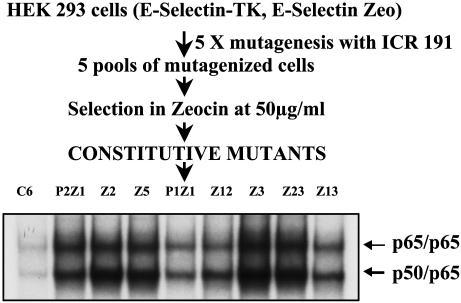
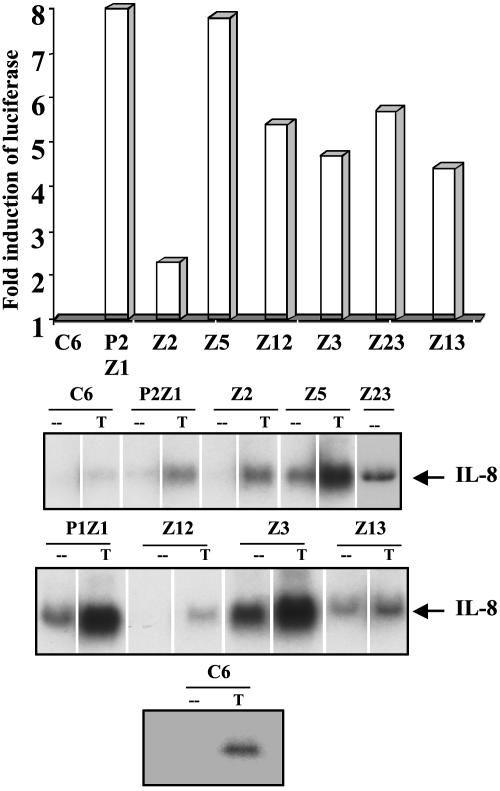
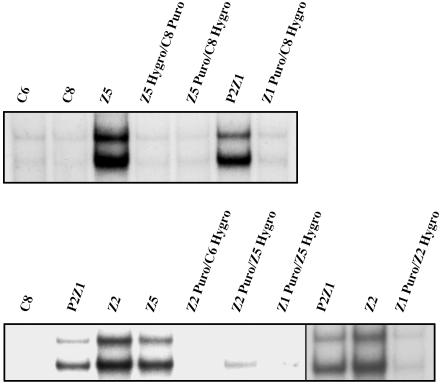
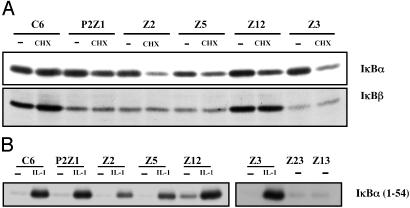
We have used a genetic approach to generate eight different mutant human cell lines in which NF-kappaB is constitutively activated. These independent clones have different phenotypes and belong to several different genetic complementation groups. In one clone inhibitor of kappaB(IkappaB) kinase is constitutively active, but in the seven others it is not, despite the fact that IkappaB is degraded in all eight clones. Thus, IkappaB kinase-independent mechanisms of IkappaB degradation and NF-kappaB activation are predominant in these mutants. Biochemical analyses of the mutants revealed that they fall into at least five different categories, differing in the sets of upstream kinases that are activated, confirming multiple mechanisms of NF-kappaB activation. By introducing a retroviral cDNA library into the Ras C6 cell line, with constitutively active NF-kappaB, followed by selection for functional complementation, we isolated a cDNA encoding a C-terminal fragment of enolase 1 and identified it as negative regulator of NF-kappaB.
Figures
Similar articles
-
Activation of NF-kappa B via the Ikappa B kinase complex is both essential and sufficient for proinflammatory gene expression in primary endothelial cells.J Biol Chem. 2001 Jul 27;276(30):28451-8. doi: 10.1074/jbc.M102698200. Epub 2001 May 3. J Biol Chem. 2001. PMID: 11337506
-
Constitutive activation of IkappaB kinase alpha and NF-kappaB in prostate cancer cells is inhibited by ibuprofen.Oncogene. 1999 Dec 2;18(51):7389-94. doi: 10.1038/sj.onc.1203160. Oncogene. 1999. PMID: 10602496
-
The role of IKK in constitutive activation of NF-kappaB transcription factor in prostate carcinoma cells.J Cell Sci. 2002 Jan 1;115(Pt 1):141-51. doi: 10.1242/jcs.115.1.141. J Cell Sci. 2002. PMID: 11801732
-
Regulation and function of IKK and IKK-related kinases.Sci STKE. 2006 Oct 17;2006(357):re13. doi: 10.1126/stke.3572006re13. Sci STKE. 2006. PMID: 17047224 Review.
-
Activation of I-kappaB kinase by the HTLV type 1 Tax protein: mechanistic insights into the adaptor function of IKKgamma.AIDS Res Hum Retroviruses. 2000 Nov 1;16(16):1591-6. doi: 10.1089/08892220050193001. AIDS Res Hum Retroviruses. 2000. PMID: 11080796 Review.
Cited by
-
Use of forward genetics to discover novel regulators of NF-kappaB.Cold Spring Harb Perspect Biol. 2010 Jun;2(6):a001966. doi: 10.1101/cshperspect.a001966. Epub 2009 Dec 23. Cold Spring Harb Perspect Biol. 2010. PMID: 20516132 Free PMC article. Review.
-
Inhibition of nuclear factor kappaB proteins-platinated DNA interactions correlates with cytotoxic effectiveness of the platinum complexes.Sci Rep. 2016 Aug 30;6:28474. doi: 10.1038/srep28474. Sci Rep. 2016. PMID: 27574114 Free PMC article.
-
Regulation of NF-kappaB by NSD1/FBXL11-dependent reversible lysine methylation of p65.Proc Natl Acad Sci U S A. 2010 Jan 5;107(1):46-51. doi: 10.1073/pnas.0912493107. Epub 2009 Dec 22. Proc Natl Acad Sci U S A. 2010. PMID: 20080798 Free PMC article.
-
The G-protein-coupled receptor kinase 5 inhibits NFkappaB transcriptional activity by inducing nuclear accumulation of IkappaB alpha.Proc Natl Acad Sci U S A. 2008 Nov 18;105(46):17818-23. doi: 10.1073/pnas.0804446105. Epub 2008 Nov 13. Proc Natl Acad Sci U S A. 2008. PMID: 19008357 Free PMC article.
-
Critical Role of Novel O-GlcNAcylation of S550 and S551 on the p65 Subunit of NF-κB in Pancreatic Cancer.Cancers (Basel). 2023 Sep 27;15(19):4742. doi: 10.3390/cancers15194742. Cancers (Basel). 2023. PMID: 37835439 Free PMC article.
References
-
- Verma, I. M., Stevenson, J. K., Schwarz, E. M., Van Antwerp, D. & Miyamoto, S. (1995) Genes Dev. 9, 2723-2735. - PubMed
-
- Liou, H. C. & Baltimore, D. (1993) Curr. Opin. Cell Biol. 5, 477-487. - PubMed
-
- Regnier, C. H., Song, H. Y., Gao, X., Goeddel, D. V., Cao, Z. & Rothe, M. (1997) Cell 90, 373-383. - PubMed
-
- DiDonato, J. A., Hayakawa, M., Rothwarf, D. M., Zandi, E. & Karin, M. (1997) Nature 388, 548-554. - PubMed
-
- Mercurio, F., Zhu, H., Murray, B. W., Shevchenko, A., Bennett, B. L., Li, J., Young, D. B., Barbosa, M., Mann, M., Manning, A., et al. (1997) Science 278, 860-866. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
