

Identification of Drosophila MicroRNA targets
- PMID: 14691535
- PMCID: PMC270017
- DOI: 10.1371/journal.pbio.0000060
Identification of Drosophila MicroRNA targets
Abstract
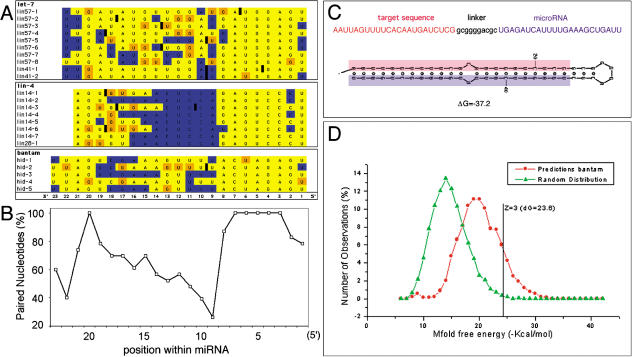
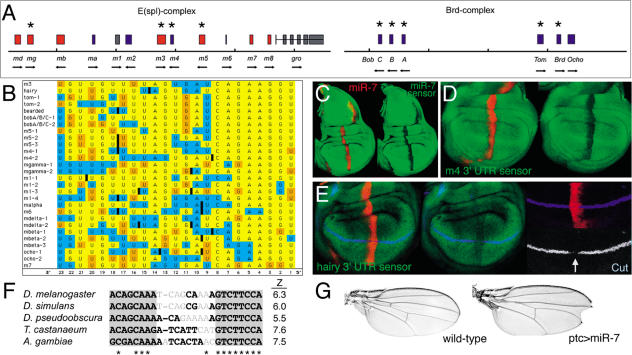
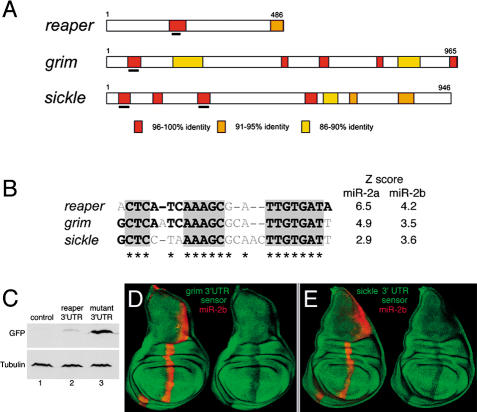
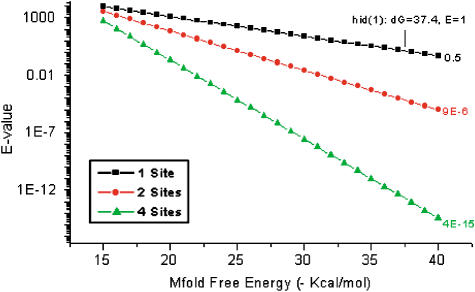
MicroRNAs (miRNAs) are short RNA molecules that regulate gene expression by binding to target messenger RNAs and by controlling protein production or causing RNA cleavage. To date, functions have been assigned to only a few of the hundreds of identified miRNAs, in part because of the difficulty in identifying their targets. The short length of miRNAs and the fact that their complementarity to target sequences is imperfect mean that target identification in animal genomes is not possible by standard sequence comparison methods. Here we screen conserved 3' UTR sequences from the Drosophila melanogaster genome for potential miRNA targets. The screening procedure combines a sequence search with an evaluation of the predicted miRNA-target heteroduplex structures and energies. We show that this approach successfully identifies the five previously validated let-7, lin-4, and bantam targets from a large database and predict new targets for Drosophila miRNAs. Our target predictions reveal striking clusters of functionally related targets among the top predictions for specific miRNAs. These include Notch target genes for miR-7, proapoptotic genes for the miR-2 family, and enzymes from a metabolic pathway for miR-277. We experimentally verified three predicted targets each for miR-7 and the miR-2 family, doubling the number of validated targets for animal miRNAs. Statistical analysis indicates that the best single predicted target sites are at the border of significance; thus, target predictions should be considered as tentative until experimentally validated. We identify features shared by all validated targets that can be used to evaluate target predictions for animal miRNAs. Our initial evaluation and experimental validation of target predictions suggest functions for two miRNAs. For others, the screen suggests plausible functions, such as a role for miR-277 as a metabolic switch controlling amino acid catabolism. Cross-genome comparison proved essential, as it allows reduction of the sequence search space. Improvements in genome annotation and increased availability of cDNA sequences from other genomes will allow more sensitive screens. An increase in the number of confirmed targets is expected to reveal general structural features that can be used to improve their detection. While the screen is likely to miss some targets, our study shows that valid targets can be identified from sequence alone.
Conflict of interest statement
The authors have declared that no conflicts of interest exist.
Figures

References
-
- Abrahante JE, Daul AL, Li M, Volk ML, Tennessen JM, et al. The Caenorhabditis elegans hunchback-like gene lin-57/hbl-1 controls developmental time and is regulated by microRNAs. Dev Cell. 2003;4:625–637. - PubMed
-
- Ambros V. MicroRNAs: Tiny regulators with great potential. Cell. 2001;107:823–826. - PubMed
-
- Ambros V, Lee RC, Lavanway A, Williams PT, Jewell D. MicroRNAs and other tiny endogenous RNAs in C. elegans. Curr Biol. 2003;13:807–818. - PubMed
-
- Aravin AA, Lagos-Quintana M, Yalcin A, Zavolan M, Marks D, et al. The small RNA profile during Drosophila melanogaster development. Dev Cell. 2003;5:337–350. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
