

Protein topology determines binding mechanism
- PMID: 14694192
- PMCID: PMC327178
- DOI: 10.1073/pnas.2534828100
Protein topology determines binding mechanism
Abstract
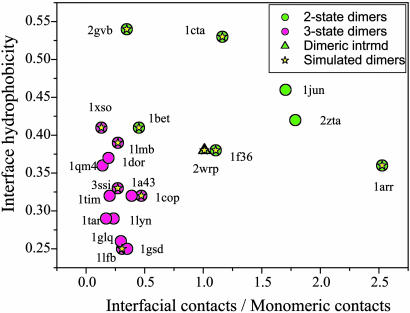
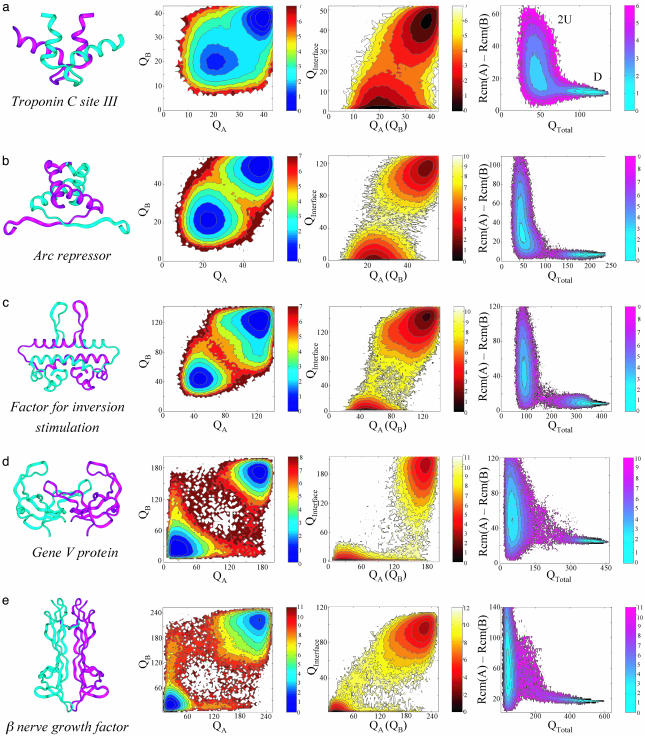
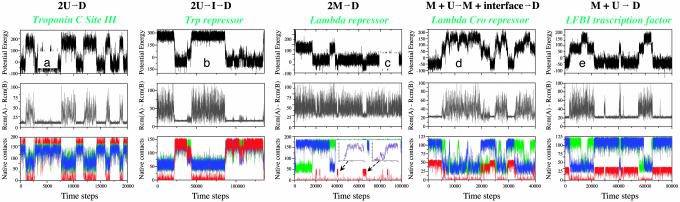
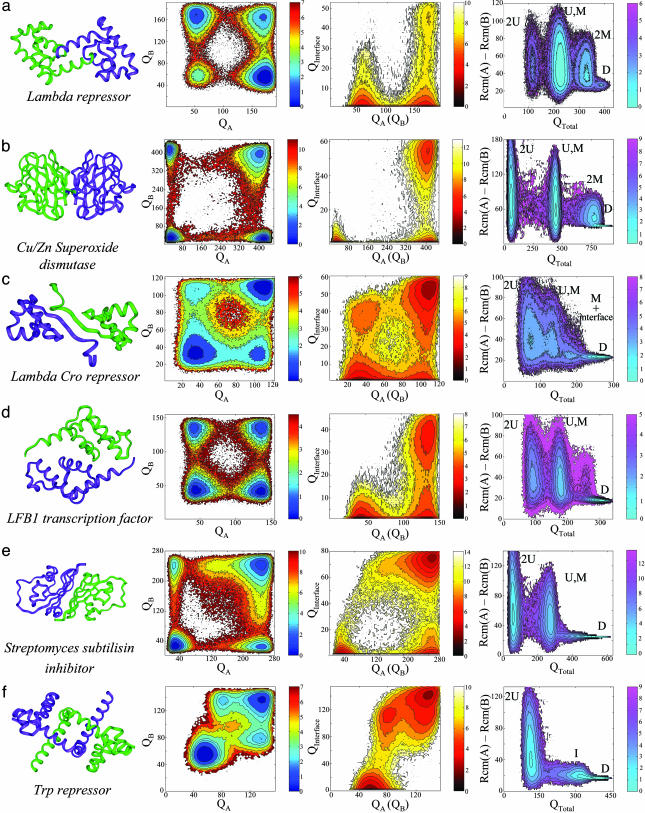
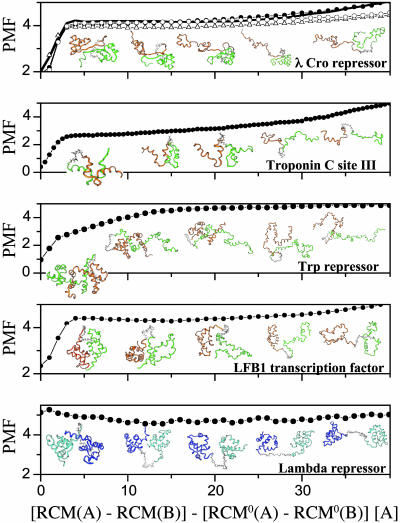
Protein recognition and binding, which result in either transient or long-lived complexes, play a fundamental role in many biological functions, but sometimes also result in pathologic aggregates. We use a simplified simulation model to survey a range of systems where two highly flexible protein chains form a homodimer. In all cases, this model, which corresponds to a perfectly funneled energy landscape for folding and binding, reproduces the macroscopic experimental observations on whether folding and binding are coupled in one step or whether intermediates occur. Owing to the minimal frustration principle, we find that, as in the case of protein folding, the native topology is the major factor that governs the choice of binding mechanism. Even when the monomer is stable on its own, binding sometimes occurs fastest through unfolded intermediates, thus showing the speedup envisioned in the fly-casting scenario for molecular recognition.
Figures
References
-
- Wodak, S. & Janin, J. (2002) Adv. Protein Chem. 61, 9–72. - PubMed
-
- Ofran, Y. & Rost, B. (2003) J. Mol. Biol. 325, 377–387. - PubMed
-
- Solomon, B., Taraboulos, A. & Katchalski-Katzir, E. (2001) Conformational Diseases—A Compendium (Bialik, Jerusalem).
-
- Onuchic, J. N., Socci, N. D., Luthey-Schulten, Z. & Wolynes, P. G. (1996) Fold. Des. 1, 441–450. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
