

Actions of a histaminergic/peptidergic projection neuron on rhythmic motor patterns in the stomatogastric nervous system of the crab Cancer borealis
- PMID: 14694531
- PMCID: PMC6494454
- DOI: 10.1002/cne.11003
Actions of a histaminergic/peptidergic projection neuron on rhythmic motor patterns in the stomatogastric nervous system of the crab Cancer borealis
Abstract
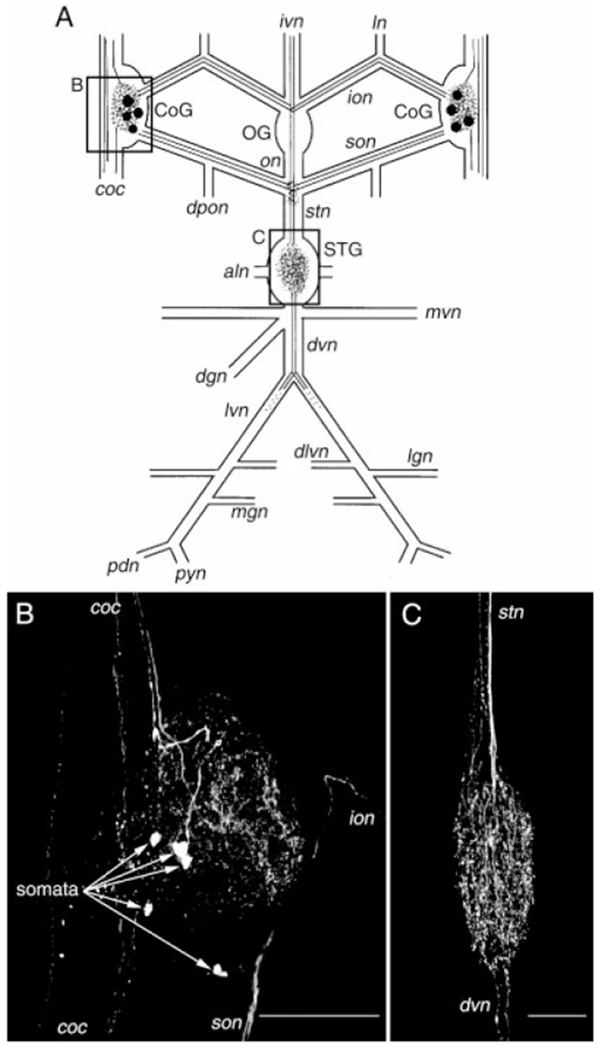
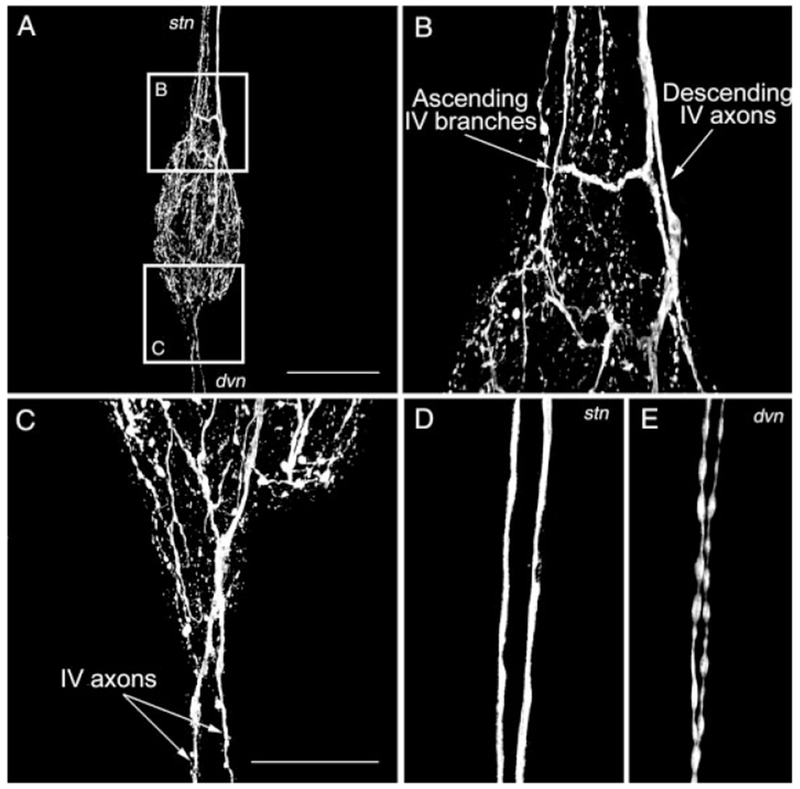
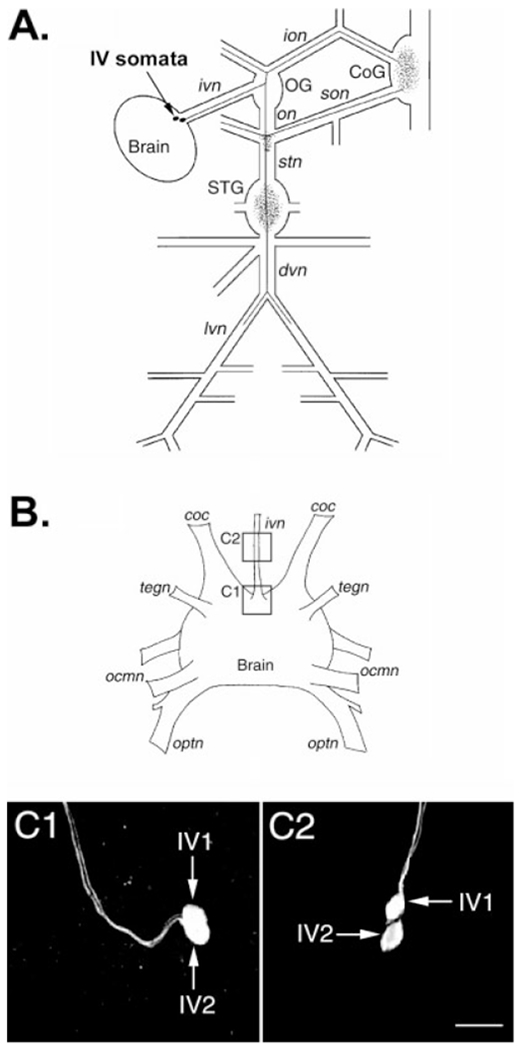
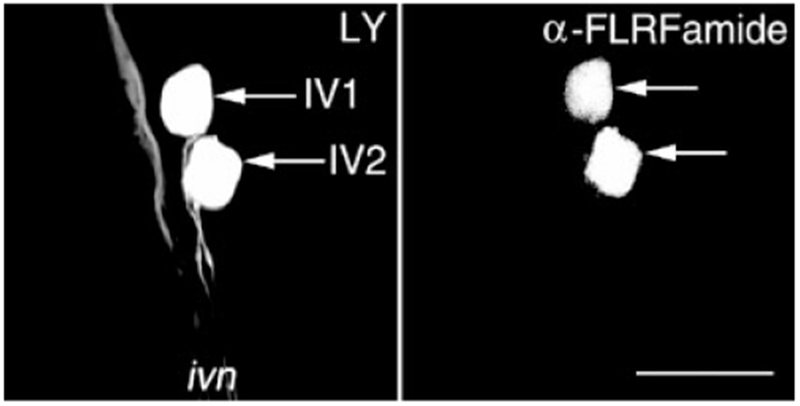
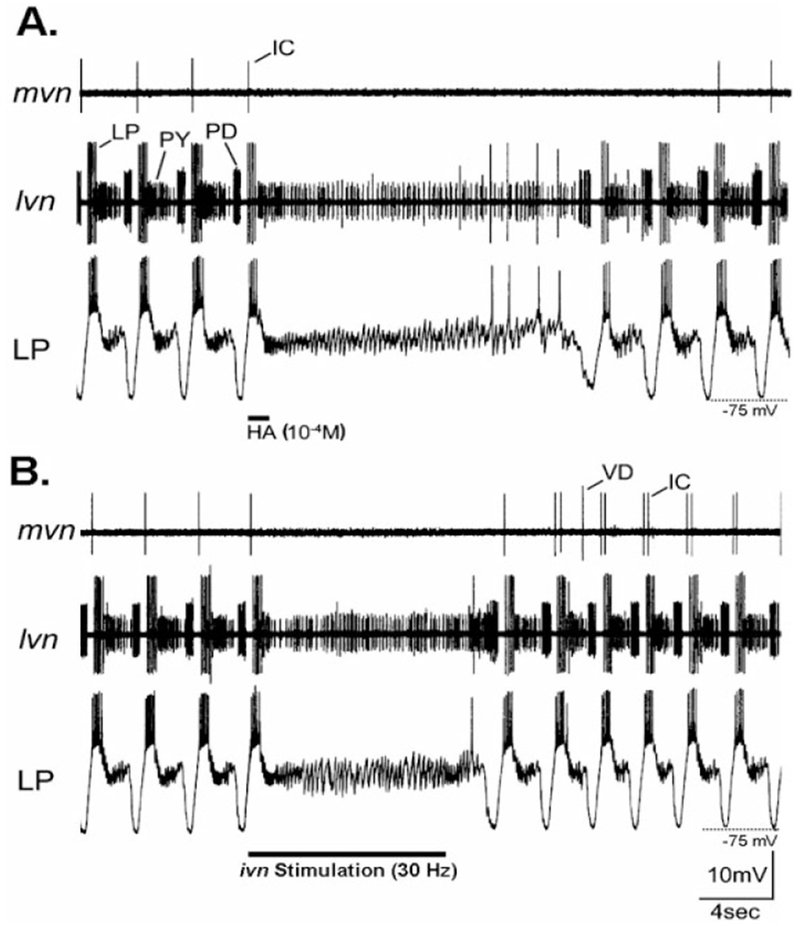
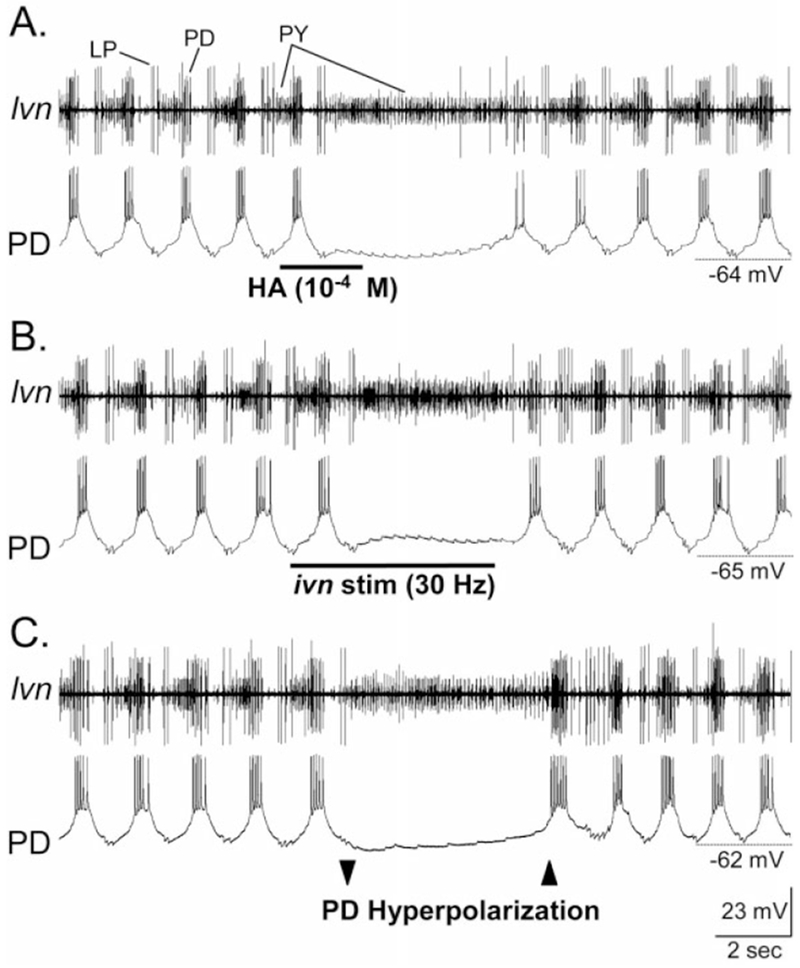
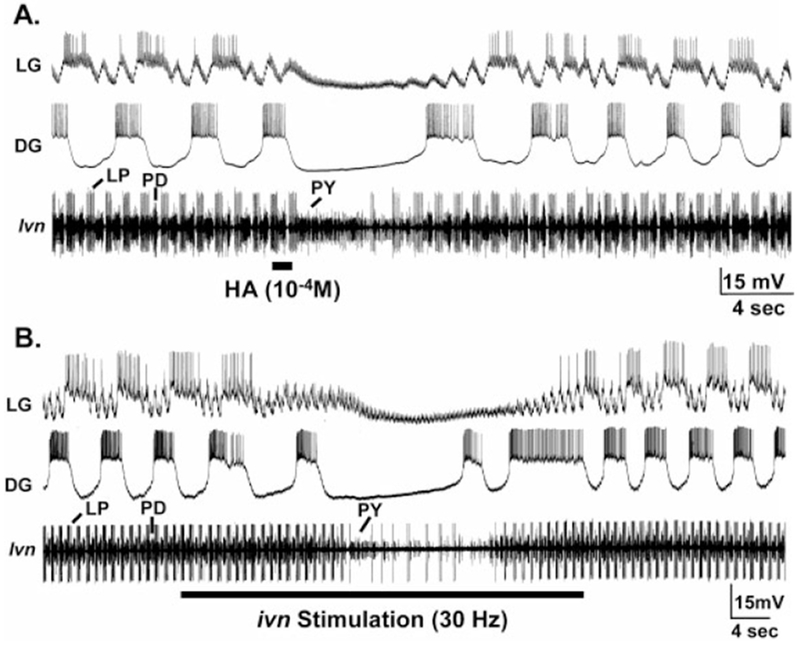
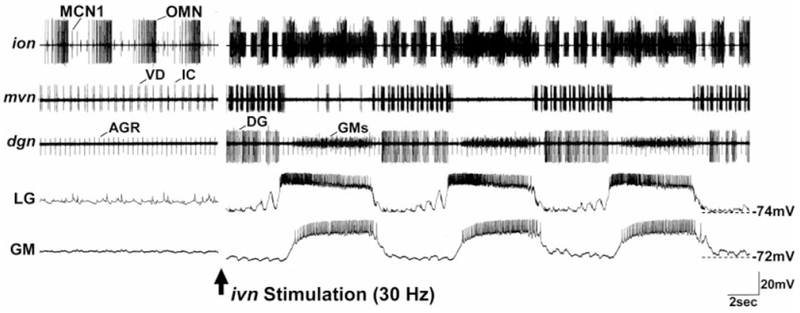
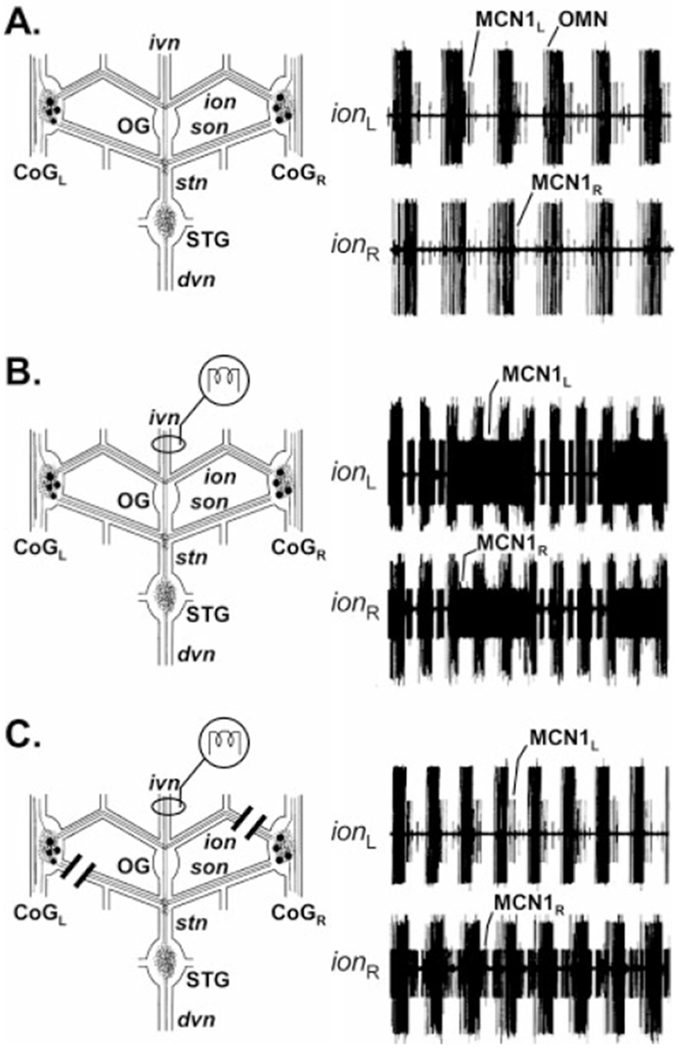
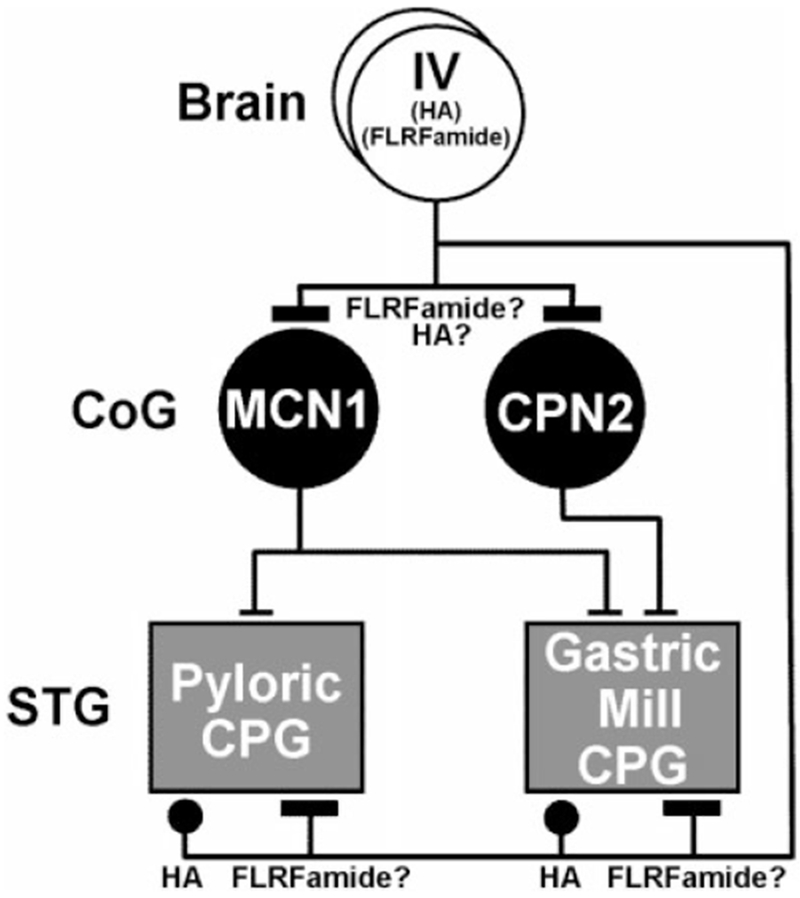
Histamine is a neurotransmitter with actions throughout the nervous system of vertebrates and invertebrates. Nevertheless, the actions of only a few identified histamine-containing neurons have been characterized. Here, we present the actions of a histaminergic projection neuron on the rhythmically active pyloric and gastric mill circuits within the stomatogastric ganglion (STG) of the crab Cancer borealis. An antiserum generated against histamine labeled profiles throughout the C. borealis stomatogastric nervous system. Labeling occurred in several somata and neuropil within the paired commissural ganglia as well as in neuropil within the STG and at the junction of the superior oesophageal and stomatogastric nerves. The source of all histamine-like immunolabeling in the STG neuropil was one pair of neuronal somata, the previously identified inferior ventricular (IV) neurons, located in the supraoesophageal ganglion. These neurons also exhibited FLRFamide-like immunoreactivity. Activation of the IV neurons in the crab inhibited some pyloric and gastric mill neurons and, with inputs from the commissural ganglia eliminated, terminated both rhythms. Focal application of histamine had comparable effects. The actions of both applied histamine and IV neuron stimulation were blocked, reversibly, by the histamine type-2 receptor antagonist cimetidine. With the commissural ganglia connected to the STG, IV neuron stimulation elicited a longer-latency activation of commissural projection neurons which in turn modified the pyloric rhythm and activated the gastric mill rhythm. These results support the hypothesis that the histaminergic/peptidergic IV neurons are projection neurons with direct and indirect actions on the STG circuits of the crab C. borealis.
Copyright 2003 Wiley-Liss, Inc.
Figures
Similar articles
-
Modulation of stomatogastric rhythms.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2009 Nov;195(11):989-1009. doi: 10.1007/s00359-009-0483-y. Epub 2009 Oct 11. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2009. PMID: 19823843 Review.
-
Projection neurons with shared cotransmitters elicit different motor patterns from the same neural circuit.J Neurosci. 2000 Dec 1;20(23):8943-53. doi: 10.1523/JNEUROSCI.20-23-08943.2000. J Neurosci. 2000. PMID: 11102505 Free PMC article.
-
Modulation of rhythmic motor activity by pyrokinin peptides.J Neurophysiol. 2007 Jan;97(1):579-95. doi: 10.1152/jn.00772.2006. Epub 2006 Oct 25. J Neurophysiol. 2007. PMID: 17065249
-
Similarities and differences in circuit responses to applied Gly1-SIFamide and peptidergic (Gly1-SIFamide) neuron stimulation.J Neurophysiol. 2019 Mar 1;121(3):950-972. doi: 10.1152/jn.00567.2018. Epub 2019 Jan 16. J Neurophysiol. 2019. PMID: 30649961 Free PMC article.
-
Frequency control of a slow oscillatory network by a fast rhythmic input: pyloric to gastric mill interactions in the crab stomatogastric nervous system.Ann N Y Acad Sci. 1998 Nov 16;860:226-38. doi: 10.1111/j.1749-6632.1998.tb09052.x. Ann N Y Acad Sci. 1998. PMID: 9928315 Review.
Cited by
-
Different sensory systems share projection neurons but elicit distinct motor patterns.J Neurosci. 2004 Dec 15;24(50):11381-90. doi: 10.1523/JNEUROSCI.3219-04.2004. J Neurosci. 2004. PMID: 15601944 Free PMC article.
-
Circuit feedback increases activity level of a circuit input through interactions with intrinsic properties.J Neurophysiol. 2017 Aug 1;118(2):949-963. doi: 10.1152/jn.00772.2016. Epub 2017 May 3. J Neurophysiol. 2017. PMID: 28469000 Free PMC article.
-
Neural circuit regulation by identified modulatory projection neurons.Front Neurosci. 2023 Mar 17;17:1154769. doi: 10.3389/fnins.2023.1154769. eCollection 2023. Front Neurosci. 2023. PMID: 37008233 Free PMC article. Review.
-
Coordination of distinct but interacting rhythmic motor programs by a modulatory projection neuron using different co-transmitters in different ganglia.J Exp Biol. 2013 May 15;216(Pt 10):1827-36. doi: 10.1242/jeb.082503. Epub 2013 Feb 7. J Exp Biol. 2013. PMID: 23393282 Free PMC article.
-
Modulation of stomatogastric rhythms.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2009 Nov;195(11):989-1009. doi: 10.1007/s00359-009-0483-y. Epub 2009 Oct 11. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2009. PMID: 19823843 Review.
References
-
- Battelle BA, Calman BG, Andrews AW, Grieco FD, Mleziva MB, Callaway JC, Stuart AE. 1991. Histamine: a putative afferent neurotransmitter in Limulus eyes. J Comp Neurol 305:527–542. - PubMed
-
- Battelle BA, Calman BG, Hart MK. 1999. Cellular distributions and functions of histamine, octopamine, and serotonin in the peripheral visual system, brain, and circumoesophageal ring of the horseshoe crab Limulus polyphemus. Microsc Res Tech 44:70–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
