

Imaging single cardiac ryanodine receptor Ca2+ fluxes in lipid bilayers
- PMID: 14695257
- PMCID: PMC1303777
- DOI: 10.1016/S0006-3495(04)74091-6
Imaging single cardiac ryanodine receptor Ca2+ fluxes in lipid bilayers
Abstract
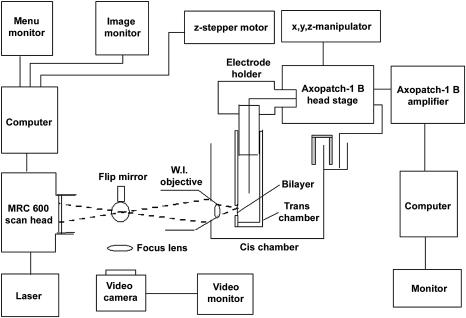
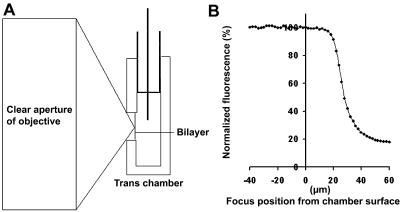
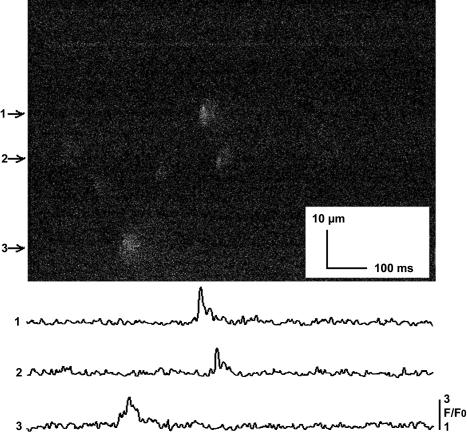
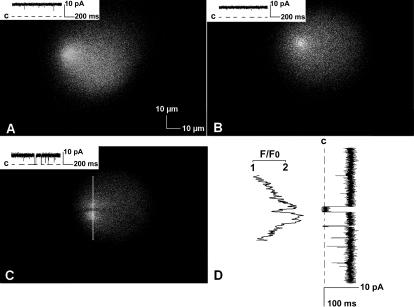
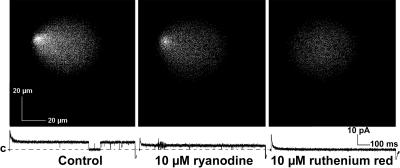
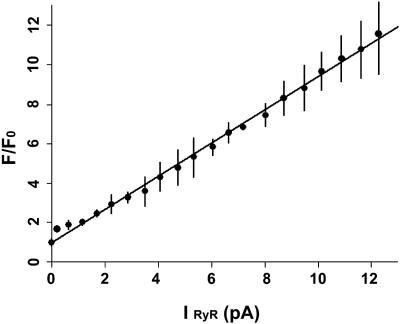
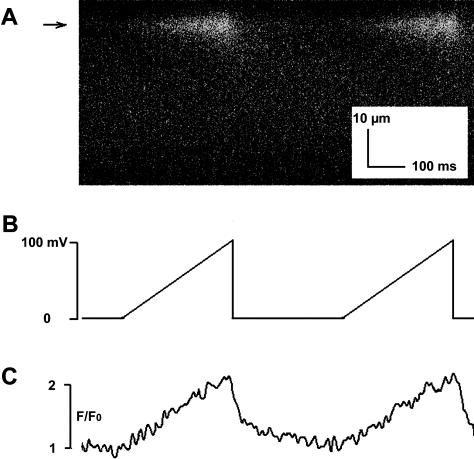
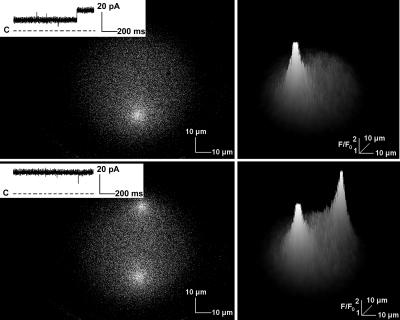
In this and an accompanying report we describe two steps, single-channel imaging and channel immobilization, necessary for using optical imaging to analyze the function of ryanodine receptor (RyR) channels reconstituted in lipid bilayers. An optical bilayer system capable of laser scanning confocal imaging of fluo-3 fluorescence due to Ca2+ flux through single RyR2 channels and simultaneous recording of single channel currents was developed. A voltage command protocol was devised in which the amplitude, time course, shape, and hence the quantity of Ca2+ flux through a single RyR2 channel is controlled solely by the voltage imposed across the bilayer. Using this system, the voltage command protocol, and concentrations of Ca2+ (25-50 mM) that result in saturating RyR2 Ca2+ currents, proportional fluo-3 fluorescence was recorded simultaneously with Ca2+ currents having amplitudes of 0.25-14 pA. Ca2+ sparks, similar to those obtained with conventional microscope-based laser scanning confocal systems, were imaged in mouse ventricular cardiomyocytes using the optical bilayer system. The utility of the optical bilayer for systematic investigation of how cellular factors extrinsic to the RyR2 channel, such as Ca2+ buffers and diffusion, alter fluo-3 fluorescent responses to RyR2 Ca2+ currents, and for addressing other current research questions is discussed.
Figures

References
-
- Airey, J. A., C. F. Beck, K. Murakami, S. J. Tanksley, T. J. Deerinck, M. H. Ellisman, and J. L. Sutko. 1990. Identification and localization of two triad junctional foot protein isoforms in mature avian fast twitch skeletal muscle. J. Biol. Chem. 265:14187–14194. - PubMed
-
- Blunck, R., J. L. Vazquez-Ibar, Y.-S. Liu, E. Perozo, and F. Bezanilla. 2003. Fluorescence measurements of KcsA channels in artificial bilayers. Biophys. J. 84:124a. (Abstr.) - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
