

The chitinolytic cascade in Vibrios is regulated by chitin oligosaccharides and a two-component chitin catabolic sensor/kinase
- PMID: 14699052
- PMCID: PMC327198
- DOI: 10.1073/pnas.0307645100
The chitinolytic cascade in Vibrios is regulated by chitin oligosaccharides and a two-component chitin catabolic sensor/kinase
Abstract
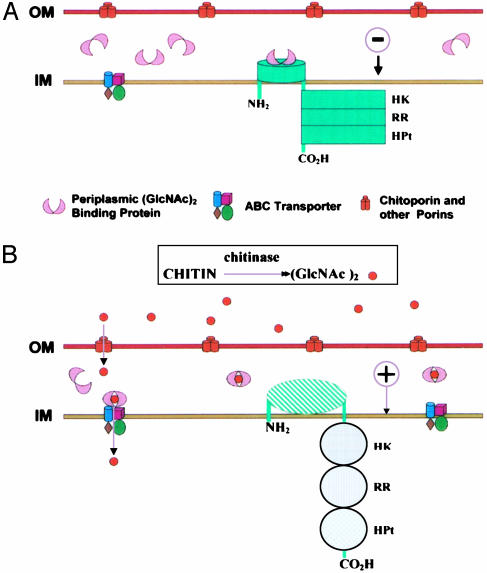
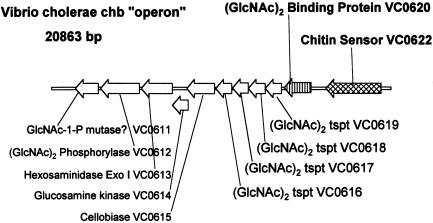
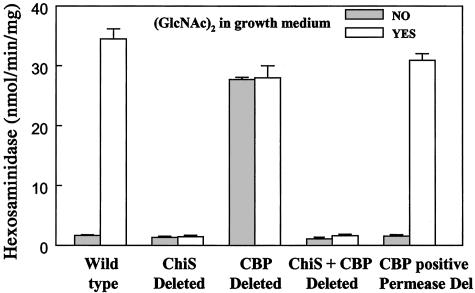
Chitin, a highly insoluble polymer of GlcNAc, is produced in massive quantities in the marine environment. Fortunately for survival of aquatic ecosystems, chitin is rapidly catabolized by marine bacteria. Here we describe a bacterial two-component hybrid sensor/kinase (of the ArcB type) that rigorously controls expression of approximately 50 genes, many involved in chitin degradation. The sensor gene, chiS, was identified in Vibrio furnissii and Vibrio cholerae (predicted amino acid sequences, full-length: 84% identical, 93% similar). Mutants of chiS grew normally on GlcNAc but did not express extracellular chitinase, a specific chitoporin, or beta-hexosaminidases, nor did they exhibit chemotaxis, transport, or growth on chitin oligosaccharides such as (GlcNAc)(2). Expression of these systems requires three components: wild-type chiS; a periplasmic high-affinity chitin oligosaccharide, (GlcNAc)(n) (n > 1), binding protein (CBP); and the environmental signal, (GlcNAc)(n). Our data are consistent with the following model. In the uninduced state, CBP binds to the periplasmic domain of ChiS and "locks" it into the minus conformation. The environmental signal, (GlcNAc)(n), dissociates the complex by binding to CBP, releasing ChiS, yielding the plus phenotype (expression of chitinolytic genes). In V. cholerae, a cluster of 10 contiguous genes (VC0620-VC0611) apparently comprise a (GlcNAc)(2) catabolic operon. CBP is encoded by the first, VC0620, whereas VC0619-VC0616 encode a (GlcNAc)(2) ABC-type permease. Regulation of chiS requires expression of CBP but not (GlcNAc)(2) transport. (GlcNAc)(n) is suggested to be essential for signaling these cells that chitin is in the microenvironment.
Figures
References
-
- Stock, A. M., Robinson, V. L. & Goudreau, P. N. (2000) Annu. Rev. Biochem. 69, 183–215. - PubMed
-
- Inouye, M. & Dutta, R. (2003) Histidine Kinases in Signal Transduction (Academic, San Diego).
-
- Keyhani, N. O. & Roseman, S. (1999) Biochim. Biophys. Acta 1473, 108–122. - PubMed
-
- Bouma, C. L. & Roseman, S. (1996) J. Biol. Chem. 271, 33457–33467. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
