

Nonmuscle myosin IIb is involved in the guidance of fibroblast migration
- PMID: 14699073
- PMCID: PMC363055
- DOI: 10.1091/mbc.e03-06-0359
Nonmuscle myosin IIb is involved in the guidance of fibroblast migration
Abstract
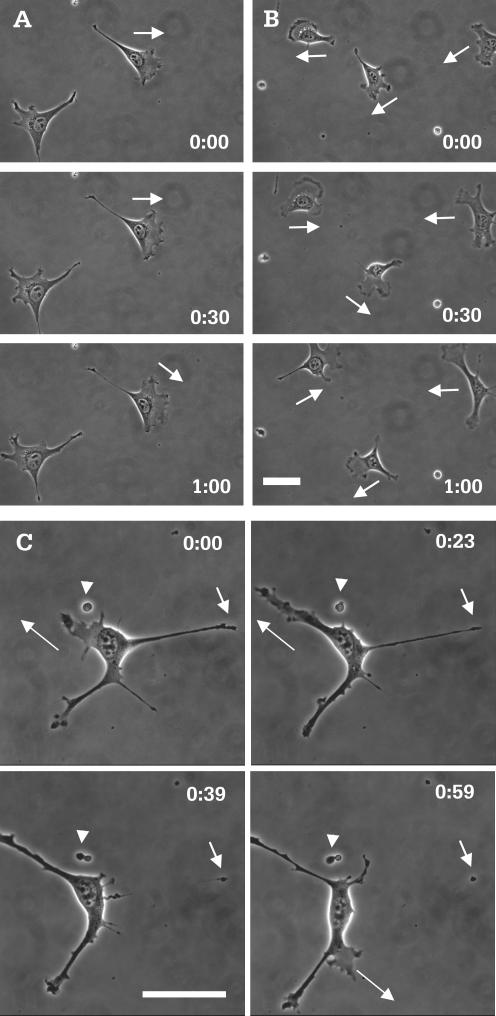
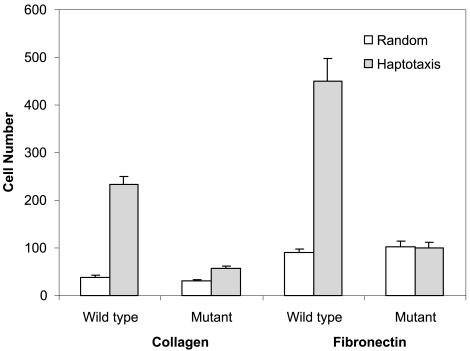
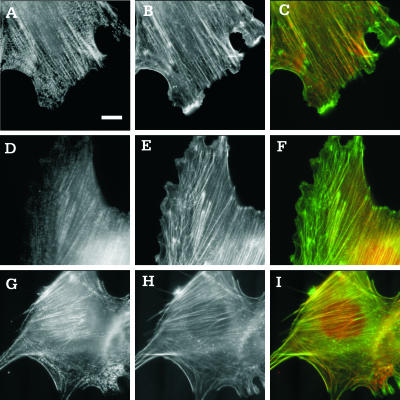
Although myosin II is known to play an important role in cell migration, little is known about its specific functions. We have addressed the function of one of the isoforms of myosin II, myosin IIB, by analyzing the movement and mechanical characteristics of fibroblasts where this protein has been ablated by gene disruption. Myosin IIB null cells displayed multiple unstable and disorganized protrusions, although they were still able to generate a large fraction of traction forces when cultured on flexible polyacrylamide substrates. However, the traction forces were highly disorganized relative to the direction of cell migration. Analysis of cell migration patterns indicated an increase in speed and decrease in persistence, which were likely responsible for the defects in directional movements as demonstrated with Boyden chambers. In addition, unlike control cells, mutant cells failed to respond to mechanical signals such as compressing forces and changes in substrate rigidity. Immunofluorescence staining indicated that myosin IIB was localized preferentially along stress fibers in the interior region of the cell. Our results suggest that myosin IIB is involved not in propelling but in directing the cell movement, by coordinating protrusive activities and stabilizing the cell polarity.
Figures





References
-
- Brown, M.E., and Bridgman, P.C. (2002). Retrograde flow rate is increased in growth cones from myosin IIB knockout mice. J. Cell Sci. 116, 1087-1094. - PubMed
-
- Burridge, K., and Chrzanowska-Wodnicka, M. (1996). Focal adhesions, contractility, and signaling. Annu. Rev. Cell. Dev. Biol. 12, 463-518. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
