

Turning heads: development of vertebrate branchiomotor neurons
- PMID: 14699587
- PMCID: PMC2219919
- DOI: 10.1002/dvdy.10444
Turning heads: development of vertebrate branchiomotor neurons
Abstract
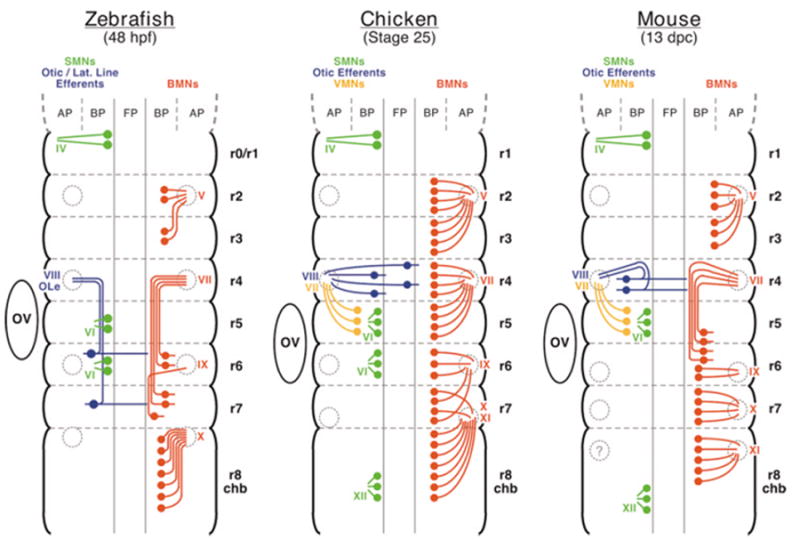
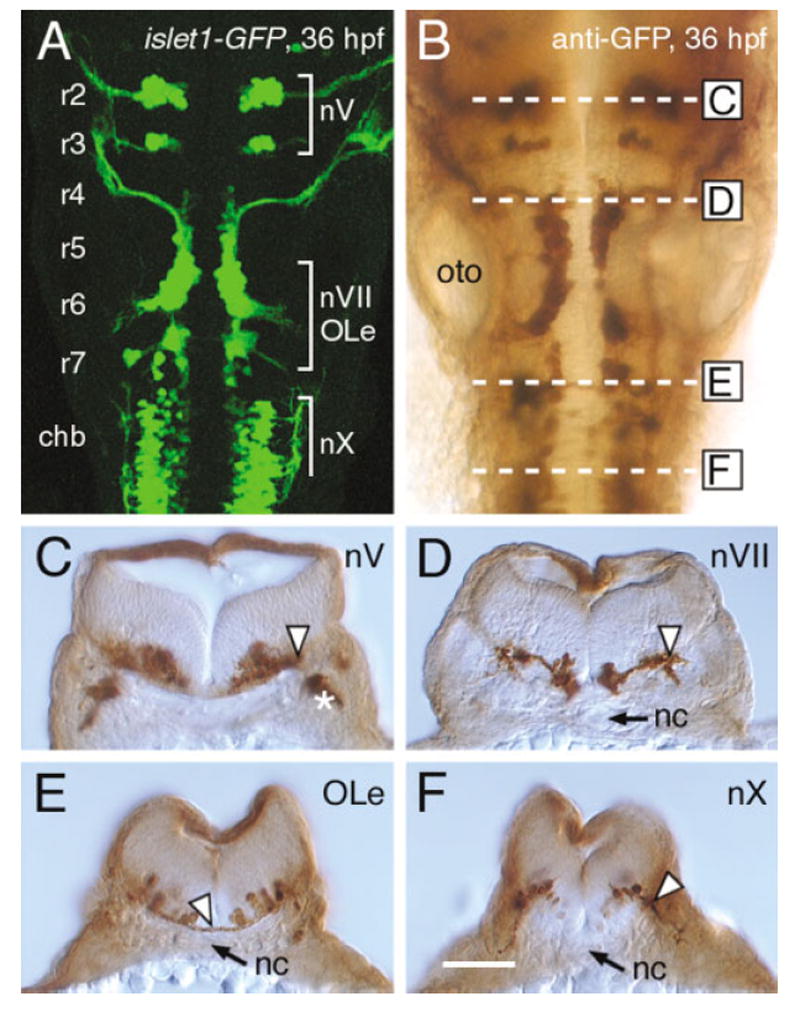
The cranial motor neurons innervate muscles that control eye, jaw, and facial movements of the vertebrate head and parasympathetic neurons that innervate certain glands and organs. These efferent neurons develop at characteristic locations in the brainstem, and their axons exit the neural tube in well-defined trajectories to innervate target tissues. This review is focused on a subset of cranial motor neurons called the branchiomotor neurons, which innervate muscles derived from the branchial (pharyngeal) arches. First, the organization of the branchiomotor pathways in zebrafish, chick, and mouse embryos will be compared, and the underlying axon guidance mechanisms will be addressed. Next, the molecular mechanisms that generate branchiomotor neurons and specify their identities will be discussed. Finally, the caudally directed or tangential migration of facial branchiomotor neurons will be examined. Given the advances in the characterization and analysis of vertebrate genomes, we can expect rapid progress in elucidating the cellular and molecular mechanisms underlying the development of these vital neuronal networks. Developmental Dynamics 229:143-161, 2004.
Copyright 2003 Wiley-Liss, Inc.
Figures
References
-
- Abadie V, Champagnat J, Fortin G. Branchiomotor activities in mouse embryo. Neuroreport. 2000;11:141–145. - PubMed
-
- Abadie V, Morisseau-Durand MP, Beyler C, Manach Y, Couly G. Brainstem dysfunction: a possible neuroembryological pathogenesis of isolated Pierre Robin sequence. Eur J Pediatr. 2002;161:275–280. - PubMed
-
- Ando H, Furuta T, Tsien RY, Okamoto H. Photo-mediated gene activation using caged RNA/DNA in zebrafish embryos. Nat Genet. 2001;28:317–325. - PubMed
-
- Appel B, Korzh V, Glasgow E, Thor S, Edlund T, Dawid IB, Eisen JS. Motoneuron fate specification revealed by patterned LIM homeobox gene expression in embryonic zebrafish. Development. 1995;121:4117–4125. - PubMed
-
- Barrett C, Guthrie S. Expression patterns of the netrin receptor UNC5H1 among developing motor neurons in the embryonic rat hindbrain. Mech Dev. 2001;106:163–166. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
